
Alinaghizadeh Rohallah
Department of Animal Genetics and Biotechnology, Agricultural College, Shahid Bahonar University of Kerman, Kerman, Iran
Mohammad Abadi Mohammadreza
Department of Animal Genetics and Biotechnology, Agricultural College, Shahid Bahonar University of Kerman, Kerman, Iran
Moradnasab Badrabadi Shahin
Department of Animal Genetics and Biotechnology, Agricultural College, Shahid Bahonar University of Kerman, Kerman, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 23 | Page No.: 4291-4294







ABSTRACT
In cattle, caseins are subdivided into four main groups: αS1-, αS2-, β- and k-caseins. k-caseins (CSN2) considerably differ from other caseins in structure and other properties. Testing the A and B alleles is of practical importance, because the milk of cows that carry the B allele of CSN3 has a better thermal resistance and shorter coagulation time, better curdles and contains micelles of different sizes. Iranian Sistani cattle (Bos indicus) are a heavy built breed and used as dual-purpose cattle breed in Eastern Iran. This breed is a genetic resource that shows special features of adaptation to rustic environments. One of the most distinctive features of Sistani cattle is its great capability to resist diseases which makes it a potential reservoir of germplasm useful for future crosses. Our main goal was to study DNA-polymorphism of the CSN3 gene in the Iranian Sistani native cattle (Bos indicus) and analyze the information value of CSN3 gene polymorphism as a genetic marker. We genotyped and analyzed 65 animals of this breed using PCR-RFLP. The frequencies of A and B alleles were 0.6385 and 0.3615 and those of AA, AB and BB genotypes were 0.4000, 0.4769 and 0.1231, respectively. In the Sistani Zebu breed, frequency of B allele is higher than other Zebu breeds, indicating that intensive selection for dairy production have been done and indirectly influenced CSN3 allele frequencies.
PDF Abstract XML References Citation
How to cite this article
Alinaghizadeh Rohallah, Mohammad Abadi Mohammadreza and Moradnasab Badrabadi Shahin, 2007. Kappa-Casein Gene Study in Iranian Sistani Cattle Breed (Bos indicus) Using PCR-RFLP. Pakistan Journal of Biological Sciences, 10: 4291-4294.
DOI: 10.3923/pjbs.2007.4291.4294
URL: https://scialert.net/abstract/?doi=pjbs.2007.4291.4294
DOI: 10.3923/pjbs.2007.4291.4294
URL: https://scialert.net/abstract/?doi=pjbs.2007.4291.4294
INTRODUCTION
Milk is the characteristic secretion of the mammary glands of all mammals. Because of its function in nutrition of the young, it is necessarily complex; it must supply nutrients, minerals and vitamins in proper form, kind and amount. Caseins are milk proteins that account for 80% of all proteins synthesized by cells of the mammary gland during lactation. In cattle, caseins are subdivided into four main groups: αS1-, αS2-, β- and κ-caseins. Casein genes of cattle are located in the q31-33 region of chromosome 6. The αS1-, αS2- and β-casein genes are linked most closely and form an evolutionarily related family, whereas the κ-casein (CSN3) gene is at least 70 kb away from others (Ferretti et al., 1990).
κ-caseins considerably differ from other caseins in structure and other properties. They are highly homologous to the γ chain of fibrinogen and fulfill a function similar to the function of this protein: they serve a stabilizing factor during the formation of micellar structure in curdling milk (Sulimova et al., 2007).
The total size of CSN3 is about 13 kb divided into 5 exons and presents two common genetic variants, A and B and these alleles differ by substitutions in 2 amino acids at positions 136 Thr(A)/Ile(B) and 148 Asp(A)/Ala(B). The variant CSN3 B is associated with processing properties, such as cheese making (Alipanah et al., 2007). Most single-nucleotide polymorphisms (SNPs) in protein-coding regions are located in exon IV. Twelve genetic variants of the CSN3 gene have been described in cattle: A, B, B2, C, E, F, F1,G, H, I, A(1) and J (Kaminski, 1996; Prinzenberg et al., 1999; Sulimova et al., 1992). One more variant (G1) has been detected in the yak (Sulimova et al., 1996). Most of these variants are rare; they are found in only a few breeds and usually at low frequencies.
The use of DNA polymorphic markers allows the determination of individual genotypes at many loci and provides information on population parameters such as allele frequencies as well as improving selection by marker assisted selection.
The SNSs in exon IV of the gene that lead to amino acid substitutions in the protein molecule also determine the appearance or disappearance of restriction sites. Several methods of typing alleles A and B of the CSN3 gene at the DNA levels have been developed to date. They use different restriction endonucleases, including HinfI, HindIII, PstI, TaqI, HaeIII and MboII (Denicourt et al., 1990; Gladyr et al., 2000; Schlieben et al., 1991; Sulimova et al., 1992, 1998). Testing the A and B alleles is of practical importance, because the milk of cows that carry the B allele of CSN3 has a better thermal resistance and shorter coagulation time, better curdles and contains micelles of different sizes, which is preferable cheese-making (Alipanah et al., 2007; Sulimova et al., 2007). The cheese yield from the milk of cows with genotype BB is 10% higher compared to the milk of AA cows (Bovenhuis et al., 1992). The B allele of CSN3 not only promotes an increase in cheese yielding capacity and improvement of cheese quality, but also is correlated with other valuable parameters of milk productivity (protein content and milk yield) (Gladyr et al., 2000; Kalashnikova et al., 2002). Selection for the B allele of CSN3 is part of cattle breeding programs in many developed countries (Pedersen, 1991). International databases contain data on CSN3 gene polymorphism in cattle breeds in 23 breeds (http://dga.jouy.inra.fr/cgiin/lgbc/Req_breeds.pl?BASE=cattle) and about 6 Russian breeds reported by Sulimova et al. (2007) and Alipanah et al. (2007) most of such studies have been done on European cattle breeds and very little information is available concerning the genetic polymorphism cattle breeds native Iran. Zakizadeh et al. (2007) studied κ-casein gene only in Iranaian Sarabi dairy cattle breed, that spreads in the Northwest of Iran. Iranian Sistani cattle are a heavy built breed and used as dual-purpose cattle breed in Eastern Iran. This black-in-color breed, that is also native to Pakistan and Afghanistan, is a genetic resource that shows special features of adaptation to rustic environments. Such characteristic has become a biotype of great interest for the meat production industry within the last few years. One of the most distinctive features of Sistani cattle is its great capability to resist diseases which makes it a potential reservoir of germplasm useful for future crosses. There are no data on Iranian Sistani cattle breed on CSN3 gene polymorphism. Our main goal was to study DNA-polymorphism of the CSN3 gene in the Iranian Sistani native cattle (Bos indicus) and analyze the information value of CSN3 gene polymorphism as a genetic marker.
MATERIALS AND METHODS
We genotyped and analyzed 65 animals of Iranian Sistani cattle breed (Bos indicus). Blood Samples (n = 65) were collected from an Iranian Sistani herd in Zehak Research Station located at Zabol (south-east of Iran) by needle punctionure of the left jugular vein, with using a 5 mL vacutainer, containing 7.5 mg EDTA. The tubes were maintained at -20°C until used for DNA extraction. Whole blood (100 μL) was used as source for DNA, which was extracted by a modified guanidine isothiocyanate- silica gel method (Boom et al., 1989). This study was conducted in Shahid Bahonar University of Kerman during autumn, 2006.
A 228 bp fragment of exon IV of the CSN3 gene was amplified by the PCR method. Briefly, the PCR was performed in a final volume of 20 μL containing 40 ng of template DNA, 10 pmol of each primer (SGE 5'-TATCATTTATGGCCATTGGACCA-3') and SGO (5'-CTTCTTTGATGTCTCCTTAGTT-3'), PCR buffer (20 mM Tris-HCl pH 8.4, 50 mM KCl), 1.5 mM MgCl2, 0.25 mM of dNTPs and 1 U of Taq DNA polymerase. DNA was denatured at 95°C for 2 min and then 35 amplification cycles were performed according to the following protocol: 30 sec at 95°C, 40 sec at 55°C and 30 sec at 72°C. The final stage of synthesis was performed at 72°C for 7 min. PCR products were electrophoresed on 1.5% agarose gels in order to check the quality and specificity of DNA fragment amplification. To examine the nucleotide sequence variability at the CSN3 locus, three restriction enzymes HinfI, HindIII and TaqI were chosen based on their ability to cut DNA in this exon. Aliquots (10 μL) of the PCR products (228 bp) were digested with the restriction enzymes according to manufacturer’s instructions.
MspI restriction fragments of plasmid pBR322 or marker M50 were used as molecular weight markers. After ethidium bromide staining, the gels were photographed under UV light with a GAS 9000 Gel Documentation System (UVItech, UK) and the relative migration of the DNA bands was estimated.
The POPGENE 3.1 (Yeh et al., 1999) and STATISTICA 6.0 (StatSoft, 1996) software packages were used for statistical analysis. To estimate the heterozygote excess in the animal samples studied, we used Selender’s coefficient: D = (Hobs –Hexp)/Hexp, where Hobs is the observed heterozygosity and Hexp is the expected heterozygosity (Pudovkin et al., 1996).
RESULTS
We selected a 228 bp segment located in exon IV of the gene for in vitro amplification to account the distribution of polymorphic restriction sites that could be used for testing the allelic variants of the CSN3 gene (Sulimova et al., 2007). This segment encodes the amino acid sequence from amino acid residue 103 to the end of the peptide and partly covers the 3'-untranslated region (primer SGO is located in the 3' region immediately behind the stop codon). It contains polymorphic restriction sites for four endonucleases: TaqI and HindIII sites of allele B and HinfI and PstI sites of allele A (Fig. 1).
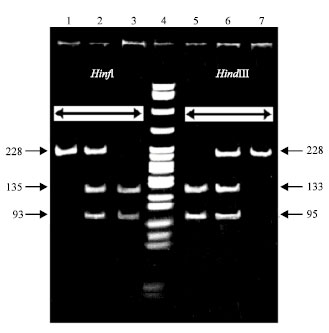 | |
| Fig. 1: | Electrophoresis of the HinfI and HindIII restriction products of DNA samples from Sistani cows. Lane 1 and 5 are BB, Lane 2 and 6 are AB, Lane 3 and 7 are AA and lane 4 is marker (MspI fragments of pBR322 plasmid DNA) |
The presence of polymorphic restriction sites for several restriction endonucleases allowed us to use any of them in subsequent experiments, which extended the possibilities of analyzing the polymorphism of the amplified fragment. The HinfI, HindIII and TaqI restrictions of amplification products were performed independently from each other in separate test tubes.
The frequencies of A and B alleles were 0.6385 and 0.3615 and those of AA, AB and BB genotypes were 0.4000, 0.4769 and 0.1231, respectively. The ratios between the observed genotype frequencies in our sample fit the Hardy-Weinberg equilibrium (G2 = 0.0419 and χ2 = 0.0417). Expected homozygosity and heterozygosity, computed using Levene (1949), Nei’s (1973) expected heterozygosity, Polymorphism Information Content (PIC) and Selender’s coefficient [D = (Hobs-Hexp)/Hexp] were 0.5348, 0.4652, 0.4617, 0.3551 and 0.005, respectively. The Iranian Sistani breed, in comparison with European cattle breeds has high frequency of the CSN3 gene B allele and BB and AB genotypes, which makes it especially valuable for dairy cattle breeding.
DISCUSSION
The method of DNA-typing of animals can be used in agricultural practice for CSN3 allele genotyping of cattle in order to reduce spreading of alleles causing low quality or quantity of production in cattle populations. This is the first study of the CSN3 gene in Iranian Sistani cattle. Also, these results will be compared with those observed in highly European Bos taurus and Bos indicus breeds. The results provided in the present study underline the high degree of polymorphism at the CSN3 gene in the Sistani cattle breed. Polymorphisms of casein genes, including the CSN3 gene are intensely studied in different breeds of cattle and these data are used as a characteristic of a breed (Gladyr et al., 2000; Sulimova et al., 2007). Frequency of B allele (0.3615) indicating that there is a tendency towards this allele. For the most Zebu breeds (Bos indicus) the B allele frequency is very low (no higher than 0.20) (Kemenes et al., 1999; Fernando et al., 2005) because these cattle have been selected for meat production not for dairy production. In the Sistani Zebu breed (Bos indicus), frequency of B allele is higher than other Zebu breeds, indicating that intensive selection for dairy production have been done and indirectly influenced CSN3 allele frequencies. Further studies should be carried out with the other bigger samples of this Zebu animals selected for meat and milk production in order to compare the allele frequencies. Selection of breeding bulls to be used at artificial insemination stations has been shown to decrease the frequency of the B allele if selection for CSN3 genotypes is not included in selection programs (Sulimova et al., 2007).
Selection for maintaining a high frequency of the B allele of the CSN3 gene in commercial cattle herds has not been performed in Iran for several years. This has resulted in a dramatic decrease in its frequency in cattle herds; in some cases, the allele has been lost altogether. It should be taken into account that the B allele of CSN3 not only promotes an increase in cheese yielding capacity and improvement of cheese quality, but also is correlated with other valuable parameters of milk productivity (protein content and milk yield) (Sulimova et al., 2007). Thus, genotyping the CSN3 gene is extremely important for selection practice, but the frequencies of its alleles cannot serve as breed-specific characteristics, because they are mainly determined by the genotypes of a limited number of breeding bulls used for artificial insemination and may be considerably changed as soon as in the next generation. In other words, CSN3 allele and genotype frequencies are unlikely to be adequate genetic markers of breed differences or phylogenetic relationships.
REFERENCES
- Boom, R., C.J. Sol, M.M. Salimans, C.L. Jansen, P.M. Wertheim-van Dillen and J. van der Noordaa, 1990. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol., 28: 495-503.
PubMedDirect Link - Ferretti, L., P. Leone and V. Sgaramella, 1990. Long range restriction analysis of the bovine casein genes. Nucl. Acids Res., 18: 6829-6833.
Direct Link - Kemenes, P.A., L.C.A. Regitano, A.J.M. Rosa, I.U. Paker and A.G. Razook et al., 1999. κ-casein, β-lactoglobulin and growth hormone allele frequencies and genetic distances in Nelore, Gyr, Guzera, Caracu, Charolais, Canchim and Santa Gertrudis cattle. Genet. Mol. Biol., 22: 539-541.
CrossRefDirect Link - Levene, H., 1949. On a matching problem in genetics. Ann. Math. Stat., 20: 91-94.
CrossRefDirect Link - Nei, M., 1973. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA., 70: 3321-3323.
PubMedDirect Link - Pudovkin, A.I., D.V. Zaykin and D. Hedgecock, 1996. On the potential for estimating the effective number of breeders from heterozygote-excess in progeny. Genetics, 144: 383-387.
PubMed - Sulimova, G.E., M. Ahani-Azari, J. Rostamzadeh, M.R. Mohammad Abadi and O.E. Lazebny, 2007. κ-casein gene (CSN3) allelic polymorphism in russian cattle breeds and its information value as a genetic marker. Russian J. Genet., 43: 73-79.
CrossRefDirect Link - Zakizadeh, S., A. Nejati-Javaremi, M. Reissmann, G. Rahimi and A. Jahan Bakhshi, 2007. Analysis of selection effect based on kappa casein gene on milk yield production of Iranian Sarabi cattle breed using stochastic simulation. Pak. J. Biol. Sci., 10: 941-945.
CrossRefPubMedDirect Link