
Hossam H. Abbas
Department of Hydrobiology,
Fagr K. Ali
Department of Water Pollution, National Research Center, Dokki, Gizza, Egypt
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 22 | Page No.: 3973-3982







ABSTRACT
In nature chromium occurs in divalent, trivalent and hexavalent forms. Hexavalent chromium predominates over the trivalent form in natural waters. Knowledge of acute toxicity of a xenobiotic often can be very helpful in predicting and preventing acute damage to aquatic life in receiving waters as well as in regulating toxic waste discharges. The 96 h LC50 tests can be used to obtain toxicity data as rapidly and inexpensively as possible. In the present study, results showed a significant decrease in total glycogen, total lipids and total protein of liver, muscles and gills after 24 and 96 h of exposure to 96 h LC50 of hexavalent chromium (43.7 mg L‾1). Cytotoxicity of metals is important because some metals are potential mutagens able to induce tumors in humans and experimental animals, the treatment of (Cr VI) at 43.7 mg L‾1 was for 24 and 96 h, respectively. DNA was extracted after treatment from brain and liver of the tested fish. Our results showed appearance of polymorphic bands at the long treatment interval (96 h) of hexavalent chromium. However, these bands were not appeared when the fish were exposed to the (Cr VI) for the short treatment interval (24 h). Histopathological changes were seen in liver, muscle and gills sections of chromium-exposed fishes. The obtained results were discussed in the study research.
PDF Abstract XML References Citation
How to cite this article
Hossam H. Abbas and Fagr K. Ali, 2007. Study the Effect of Hexavalent Chromium on Some Biochemical, Citotoxicological and Histopathological Aspects of the Orechromis spp. Fish. Pakistan Journal of Biological Sciences, 10: 3973-3982.
DOI: 10.3923/pjbs.2007.3973.3982
URL: https://scialert.net/abstract/?doi=pjbs.2007.3973.3982
DOI: 10.3923/pjbs.2007.3973.3982
URL: https://scialert.net/abstract/?doi=pjbs.2007.3973.3982
INTRODUCTION
Pollution of water resources is a serious and growing problem but despite the existence of relevant legislation the pollution of the aquatic environment by toxic chemical pollutants continues to occur, with domestic and industrial effluents being the main sources responsible for the contamination of aquatic environments (Claxton et al., 1998; White and Rasmussen, 1998). Contamination of the environment by heavy metals has been increasing every year (Majer et al., 2002) and the analysis of the cytotoxic effects of such metals has received special attention due to the fact that they are potentially mutagenic and induce the formation of tumors in experimental organisms and humans exposed to them (Garcia-Rodríguez et al., 2001).
Heavy metals due to their bio-accumulative and non-biodegradable properties constitute a core group of aquatic pollutants. Chromium particulates enter the aquatic medium through effluents discharged from tanneries, textiles, electroplating, metal finishing, mining, dyeing and printing industries, ceramic, photographic and pharmaceutical industries etc. They concentrate in the tissues of aquatic biota and are known to produce cumulative deleterious effects (Abbas, 1998; Abbas and Mahmoud, 2003, 2004).
Chromium was proved to be highly toxic to Oreochromis sp. and the harmful effects caused by chromium to the fish include depression in metabolic rate, histopathological and hematological alterations (Sesha and Balaparameswara, 1996, 1998, 1999), a number of studies have suggested that chromium is an essential nutrient for humans and animals (Pan et al., 2003). Chromium appears to play an important role in carbohydrate metabolism. A perusal of the available literature reveals that studies on the acute effects of hexavalent Chromium on the biochemical constituents and histopathological changes of fishes are scanty (Sridevi and Reddy, 2000; Gatta et al., 2001; Negilski, 2002).
Hexavalent chromium potentially possesses toxicological, carcinogenic and mutagenic properties (Langard and Norseth, 1979). It concentrated in the tissues of fish enter human beings through food chain and due to their cumulative action causes potential health hazards sometimes even lethal (Abbas et al., 2007). The toxic effects may result from the bioconcentration of metals and their consequent binding with biologically active constituents of the body such as lipids, amino acids, enzymes and proteins (Passow et al., 1961). It can damage DNA in several ways, including DNA Double Strand Breaks (DSBs) which generate chromosomal aberrations, micronucleus formation, sister chromatid exchange, formation of DNA adducts and alterations in DNA replication and transcription. We have attempted to characterize the effects of (Cr VI) on DNA damage in the most popular fish in Egypt like Nile tilapia (Oreochromis niloticus).
Chromium compounds are known to have toxic, genotoxic, mutagenic and carcinogenic effects on man and animals (Von Burg and Liu, 1993; Stohs and Bagchi, 1995; Mount and Hockett, 2000). Trivalent chromium III (Cr III) and hexavalent chromium VI (Cr VI) are being biologically active but differing in their ability to cross biological membranes. Octahedral chromium III is potentially mutagenic but does not pose an immediate danger to cells because it is unable to cross cell membranes, although it may be environmentally transformed into tetrahedral hexavalent chromium VI (chromate) to which cell membranes are highly permeable (Leonard and Lauwerys, 1980; Beyersmann et al., 1994; Stohs and Bagchi, 1995; Singh et al., 1998; Matsumoto, 2003).
Several in vivo and in vitro studies have shown that chromium compounds damage DNA in a variety of ways, including DNA Single and Double-Strand Breaks (SDSBs) generating chromosomal aberrations, micronucleus formation, sister chromatid exchanges, formation of DNA adducts and alteration in DNA replication and transcription (O'Brien et al., 2001; Matsumoto, 2003; Matsumoto and Marin-Morales, 2004).
Organisms used in mutagenesis testing should be selected using criteria that permit a realistic evaluation of the potential of a suspected mutagen to induce changes in genetic material. Aquatic organisms such as fish accumulate pollutants directly from contaminated water or indirectly through the ingestion of contaminated aquatic organisms. Fish are used as a test organism in which it is possible to detect DNA damage induced by direct mutagens and pro-mutagens in both fresh and salt water (Mitchelmore and Chipman, 1998; Lemos et al., 2005). Thus, genotoxic pollutants may lead to the contamination not only of the aquatic organisms themselves but of the entire ecosystem and, finally, of humans through the food chain.
In Egypt, the effect of heavy metals on fishes has been of great importance since fish is becoming a necessity in view of the high demand for fish as a relatively cheap source of animal protein and may compensate for the present deficiency of other expensive proteins.
Fish are often at the top of the aquatic food chain and may concentrate large amounts of some metals such as chromium, this metal accumulate in fish organs and made many disturbances in its vital processes (Gomaa et al., 1995). In this context, all attempt was made to investigate the acute effects of hexavalent chromium on some biochemical profiles of the tilapia fish; Oreochromis sp. since it is a widely cultured edible fish in Egypt and also forms an important link in the aquatic food chain. Biochemical profiles in fish and other aquatic organisms under heavy metal stress serve as important bioindicators in the monitoring of aquatic environment (Abbas and Mahmoud, 2004; Shalaby et al., 2005; Abbas, 2006; Abbas et al., 2007).
Therefore, the main objective in the present study is to assess the biochemical and histopathological changes and to evaluate the genotoxicty potential of hexavalent chromium on tilapia fish (Oreochromis sp.).
MATERIALS AND METHODS
Oreochromis sp. were obtained from a private sector fish farm at Egypt-Alexandria desert road 65 km from Cairo in June, 2006 and healthy fish were acclimated in the laboratory for 7 days in glass aquaria of 63 L capacity (35χ60χ30 cm) containing well-aerated unchlorinated tap water before they were used for experiments. Feeding was done once daily using a pelleted diet at rate 3% of body weight of the fish. Feeding was discontinued 48 h prior to and during the study. The water in aquaria was changed daily to avoid fish metabolite accumulations in glass aquaria. The water in aquaria was aerated 24 h before use and then aeration ceased while experimentation. Water characteristics used were measured according to the method of APHA (1995) (Table 1). Biochemical analysis was carried out on fingerlings of Oreochromis sp. (total length 7.8-9.3 cm and total weight 8.5-11.4 g). Twenty fish in 2 batches of ten each were exposed separately for 24 and 96 h to the 96 h LC50 concentration of chromium (43.7 mg L‾1) which was determined earlier according to Behreues and Karbeur (1953). Five fish were sacrificed both at the end of 24 h and also at 96 h, blotted dry and weighed. They were later dissected to isolate the whole liver, muscle and whole gill tissues. The tissues were dried for 24 h in a hot air oven at 50°C. The dry tissues were weighed to the nearest mg. The tissues of the control fishes (n = 5) were also processed similarly for biochemical analysis. The biochemical parameters, glycogen, total lipid and total protein were analyzed by adopting standard protocols (Handle and Van, 1965; Schmit, 1964; Henry, 1964) (Table 2, 3), respectively. The biochemical constituents were expressed as mg per gram dry weight of the tissue and only the arithmetic mean values (n = 5) are presented to express the results. The mean values of the control and the exposed fishes are compared following t-test (SAS, 2000).
| Table 1: | Characteristics of the test water used in the experiment |
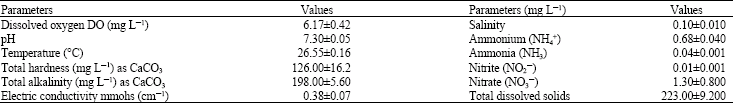 | |
| Data are presented as mean of three samples±SEM | |
| Table 2: | Effect of 96 h LC50 concentration of hexavalent chromium (43.7 mg L‾1) on total glycogen, total lipids and total protein concentration of liver, muscle and gill of Oreochromis sp. (mg dry‾1 weight‾1) at the end of 24 h exposure (n = 5) |
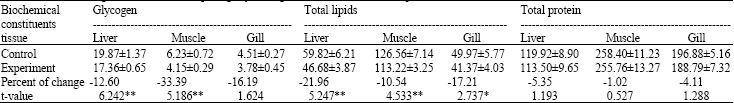 | |
| Data are represented as Mean±Standard error. *: Significant; **: Highly significant | |
| Table 3: | Effect of 96 h LC50 concentration of hexavalent chromium (43.7 mg L‾1) on total glycogen, total lipids and total protein concentration of liver, muscle and gill of Oreochromis sp. (mg dry‾1 weight) at the end of 96 h exposure (n = 5) |
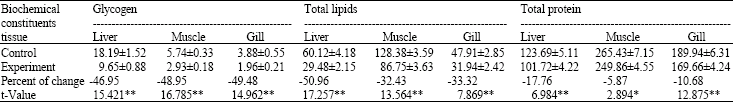 | |
| Data are represented as Mean±Standard error; *: Significant; **: Highly significant | |
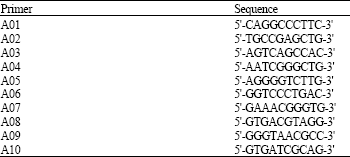
| Table 4: | Sequence of primers employed |
 | |
Molecular analysis
DNA extraction: The genomic DNA was isolated from brain and liver of the fish using phenol/chloroform extraction and ethanol precipitation method with minor modifications (Sambrook et al., 1989). The purity of the DNA preparation was judged by examining the ratio of absorbency at 260-280 nm (Aquardo et al., 1992).
RAPD-PCR analysis: To generate RAPD profiles from tilapia DNA, 21 oligodecamers (10 mer random primers: A01, A02, A03, A04, A05, A06, A07, A08, A09 and A10 (Table 4) from the Operon Technologies were used. DNA amplification reactions were performed under conditions reported by Williams et al. (1990) and Plotsky et al. (1995). PCR amplification was conducted in 20 μL reaction volume containing 100 ng genomic DNA; 100 μM dNTPs; 40 nM primer (Operon, Almeda, CA, USA); 2.5 units of Taq DNA polymearse and 5 μL promega 10X Taq DNA polymearse buffer. The reactions were carried out in Thermocycler (Perkin-Elmer 9700) programmed with a first denaturation of 5 min at 94°C, followed by 45 cycles of 1 min at 94°C, 1 min at 36°C and 2 min at 72°C and finally, one cycle at 72°C for 5 min. The PCR product was analyzed by electrophoresing 15 μL of the amplified mixture on agarose gel. The Gel-Pro Analyzer (Media Cybernetics) was used to document ethidium bromide DNA gels.
Histopathological study: Tissue specimens from liver, muscle and gills were taken from tilapias that were exposed to hexavalent chromium by the end of 96 h of exposure. The specimens were fixed in 10% buffered formalin then later processed by conventional method, sectioned at 4 μm and stained with Haematoxylin and Eosin (Bancroft et al., 1996).
RESULTS AND DISCUSSION
The glycogen, total lipid and total protein levels in liver, muscle and gill of control fish and of Oreochromis sp. exposed to the 96 h LC50 concentration of chromium for 24 and 96 h were presented (Table 2, 3). It is clear from the results that there is a decline in different biochemical constituents. The glycogen concentration of the liver, muscle and gill of the exposed fishes was 17.36, 4.15 and 3.78 mg g‾1 dry weight, respectively, showing a decrease of 12.6, 33.39 and 16.19% from control at the end of 24 h exposure (Table 2) and 9.65, 2.93 and 1.96 mg g‾1 dry weight showing a decrease of 46.95, 48.95 and 49.48% from control at the end of 96 h, respectively (Table 3).
The results of the t-test show that the decrease in the glycogen level from control is significant in liver and muscle except in gill at the end of 24 h. This decrease was, however, highly significant in all three tissues at the end of 96 h exposure.
The total lipid concentration of liver, muscle and gill of the exposed fishes was 46.68, 113.22 and 41.37 mg g‾1 dry weight (Table, 2) of the tissue showing a significant decrease of 21.96, 10.54 and 17.21% from control at the end of 24 h exposure and 29.48, 86.75 and 31.94 mg g‾1 dry weight (Table 3) showing a significant decrease of 50.96, 32.43 and 33.32% from control at the end of 96 h, respectively.
The total protein concentration of liver, muscle and gill was 113.50, 255.76 and 188.79 mg g‾1 dry weight, respectively and it decreased by 5.35, 1.02 and 4.11% from control, respectively, in fishes exposed for 24 h (Table 2) and 101.72, 249.86 and 169.66 mg g‾1 dry weight of the tissue showing a significant decrease of 17.76, 5.87 and 10.68% (Table 3) from control at the end of 96 h exposure, respectively.
The results of the present study indicated that the glycogen, total lipid and total protein concentration of liver, muscle and gill were depleted in Oreochromis sp. exposed to 96 h LC50 concentration of hexavalent chromium. A decrease in the glycogen content of fishes exposed to metallic stress was observed by many investigators (Quyyam and Shaffi, 1977; Gill and Pant, 1981; Radhakrishnaiah el al., 1992; James et al., 1995; Mary and Reddy, 1996; El-Nagar et al., 2001; Abbas et al., 2002; Abbas and Mahmoud, 2003, 2004; Mahmoud and Abbas, 2004; Shalaby et al., 2005; Abbas, 2006).
The decrease in the glycogen concentration of the tissues of Oreochromis sp. may be due to its enhanced utilization since glycogen forms the immediate source of energy to meet energy demands under metallic stress. DeZwaan and Zandee (1973) stated that decrease in glycogen concentration might be due to the prevalence of hypoxic or anoxic conditions, which normally enhances glycogen utilization. Enhanced utilization of glycogen and its consequent depletion in tissues may be attributed to hypoxia since it increases carbohydrate consumption. Under hypoxic conditions, the animal derives its energy from anaerobic breakdown of glucose, which is available to the cells by the increased glycogenolysis (Mary and Reddy, 1996; El-Nagar et al., 2001; Abbas, 2006). Present studies on the respiration of Oreochromis sp. exposed to chromium have also shown decreased oxygen consumption indicating hypoxia. Behavioral manifestations of acute toxicity like copious mucous secretion, surfacing and darting movements, darkening of skin on dorsal side, loss of scales and loss of equilibrium were observed in Oreochromis sp. under chromium stress (Sesha and Balaparameswara, 1996, 1999). Chromium induced hypoxia probably might have resulted in a shift to anaerobic glycolytic pathway by increased glycogenolysis. Depleted glycogen levels following chromium stress reported in Cyprinus carpio communis (Ambrose et al., 1994; Shalaby et al., 2005) under hypoxic conditions also supports this view. A consistent decrease in tissue glycogen reserves observed in this study suggests impaired glycogenesis. Further, the decline in glycogen might be partly due to its utilization in the formation of glycoproteins and glycolipids, which are essential constituents of various cells and other membranes.
Decrease in tissue lipid and proteins were also observed in Labeo rohita exposed to chromium. Earlier studies have also shown that lipid and protein concentration of vital organs like gills, liver, muscle and kidney depleted in fishes exposed to chromium (Arillo et al., 1982; Sastry and Sunitha, 1984; Ambrose et al., 1994). The decrease in tissue lipid and proteins might be partly due to their utilization in cell repair and tissue organization with the formation of lipoproteins which are important cellular constituents of cell membranes and cell organelles present in cytoplasm (Harper, 1963). Damage to the gill tissue was also observed in Labeo rohita exposed to chromium (Sesha and Balaparameswara, 1998) and the decrease in the lipid concentration observed in the present study can also be attributed to its utilization in cell repair and tissue organization.
The depletion in tissue proteins of Oreochromis sp. may be due to impaired or low rate of protein synthesis under metallic stress as has been reported earlier (Syversen, 1990; Prasanta and Milan, 1996; El-Nagar et al., 2001; Abbas et al., 2002; Abbas and Mahmoud, 2003, 2004; Mahmoud and Abbas, 2004; Shalaby et al., 2005; Abbas, 2006) or due to their utilization in the formation of mucoproteins which are eliminated in the form of mucous. Further, direct and/or indirect utilization of proteins and lipids for energy needs was also reported (Nagai and lkeda, 1971). Also, the utilization of proteins in cell repair and organization as causes of their depletion in the tissues cannot be ruled out. The present study showed that chromium is toxic to Oreochromis sp. and induced alterations at the biochemical level, more pronounced changes occurring at the end of 96 h and thus it is time-dependent. Also, the metal induced alterations may probably effect the enzyme mediated biodefence mechanisms of the fish. Further studies are required to elucidate the impact of chromium on detoxifying enzymes for assessing the fish health.
RAPD fingerprinting pattern: The genomic density of the fingerprints generated by the arbitrary primers used was evaluated among tilapia fish. All of primers used gave positive and detectable bands (Fig. 1). These random primers amplified a total of 108 different bands, ranging from 136 to 1305 base pairs. Over all samples, the number of RAPD bands generated per primer varied between 8 and 20 bands, with an average 11.3±0.9 bands per primer. Nearly the same results were obtained when the PCR assay was performed for each sample within each group (5 fish).
Of the scorable bands, 43 (40%) were similar monomorphic for control and treated brain and liver fish samples. Most these bands were monomorphic for control and short duration treated fish (Fig. 1). However, the DNA of the brain and liver samples treated with chromium for long time revealed the appearance of 65 (60%) new bands (polymorphic), which did not appear in the samples of other groups (Fig. 1). These new bands could be considered as genus diagnostic markers which attributed to long duration of chromium treatment. In addition, the results showed that most primers used (P01, P02, P03, P04, P06, P07 and P10) appeared polymorphic bands in the liver at the long time intervals than in the brain (Fig. 1).
As known, chromium stably exists in the environment as Cr (III) or Cr (VI). Cr (VI) is more toxic because it is bioavailable and produces radicals during reduction to Cr (III). In the present study, Nile tilapia was exposed to (Cr VI) to determine the genotoxicity using the RAPD-PCR. We have found that (Cr VI) was more toxic at the long time interval than short treatment interval. Where, it was able to creates DNA adducts (polymorphic bands) at the long time interval in brain and liver of the Nile tilapia. The appearance rate of these DNA adducts was low at the short time interval. Present results agree with those reported by Von Burg and Liu (1993), Blasiak and Kowalik (2000), Matsumoto (2003) and Matsumoto et al. (2006), who proposed that chromium exerts a genotoxic effect on animals due to its potential to cause various forms of DNA damage Calamari et al. (1982) found that the genotoxic effect of the chromium was clearer in the liver than other organs, when the adult males of Salmo gairdneri fish were exposed to Cromium at 32 mg L‾1. These findings support our results, because the polymorphic band patterns produced from Cr(VI) treatment were more pronounced in the liver than in the brain of the Nile tilapia. Calamari et al. (1982) also suggested that chromium bioaccumulates in the liver more than other organs.
The action mechanism of Cr (VI) to induce the genotoxicity it is not clearly understood. According to Sugiyama (1992), hexavalent chromate enters cells via the surface transport system and inside the cell it is reduced to trivalent chromium which induces genotoxic effects in the cell (Bianchi et al., 1983). However, if trivalent chromium has access to the intracellular medium through processes such as pinocytosis and endocytosis or by the reduction of hexavalent chromium inside the cell it acts directly on DNA and causes more damage than when it continues in the chromate form (Matsumoto, 2003). In addition, Dayan and Paine (2001) suggested that the Cromium binds macromolecules, forms adducts with thiol groups on proteins, creates DNA adducts and causes DNA strand breakage.
Recently, Roling and Baldwin (2006) found that Cromium is able to alter negatively the expression of several genes in Fundulus heteroclitus adult male livers including aldehyde dehydrogenase 4 (ALDH4), several glutathione S-transferases (GSTs) and an EST. ALDH4 and the GSTs protect the cell from cellular and oxidative stress (Hartley et al., 1995; Roling and Baldwin, 2006).
The above biochemical investigations were confirmed by the histological sections. The liver is generally regarded as central organ of metabolism in fish; structural damages from the effects of chromium in liver metabolism have been supported by the results of biochemical studies, which provide early warning indicators of toxicological responses. In addition, the liver of the treated fish showed vacuolar degeneration with activation of melanomacrophage centers (Fig. 2A) and diffuse vacuolar degeneration with congestion and dilation of sinusoids (Fig. 2B).
The vacuolization of hepatocytes might indicate an imbalance between the rate of synthesis of substances in the parenchymal cells and the rate of their release into the circulation system (Gingerich, 1982). Because the excretion of the divalent ions is a major function of the renal tubular epithelium, pollution with chromium would be highly likely to affect these cells. The negative effect of chromium in the liver exposed has been suggested to be an indicator of liver dysfunction for a variety of chemicals (Jiraungkoorskul et al., 2003).
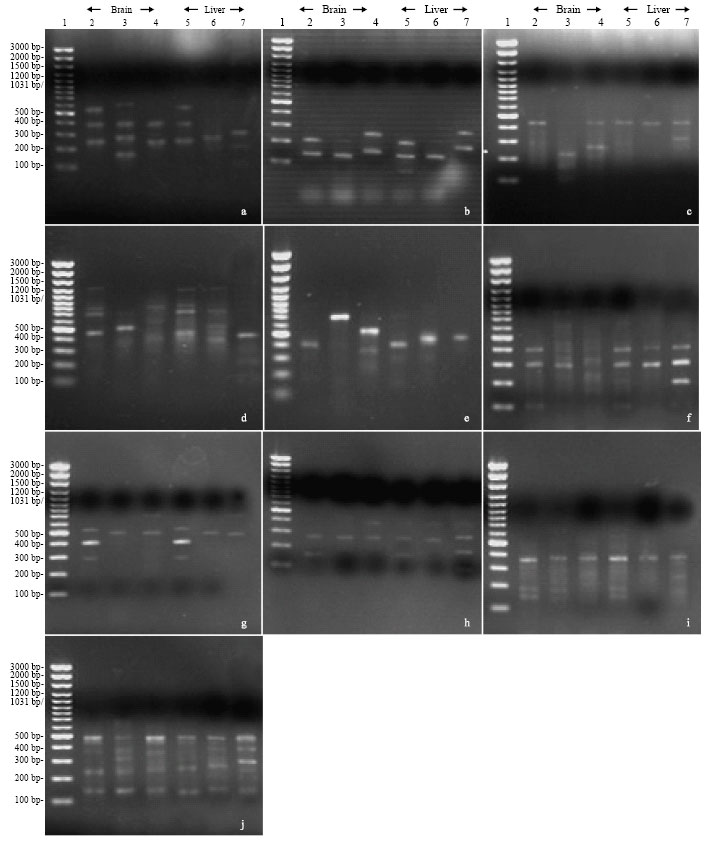 | |
| Fig. 1: | Comparison of RAPD fingerprinting profiles of different tilapia genomic DNA. a, b, c, d, e, f, g, h, i and j agarose gels represent PCR products with primer A01, A02, A03, A04, A05, A06, A07, A08, A09 and A10, respectively. Lane 1 represents DNA marker. Lanes 2 and 5 represent untreated fish. Lanes 3 and 6 represent fish treated with chromium for 24 h. Lanes 4 and 7 represent fish treated with chromium for 96 h |
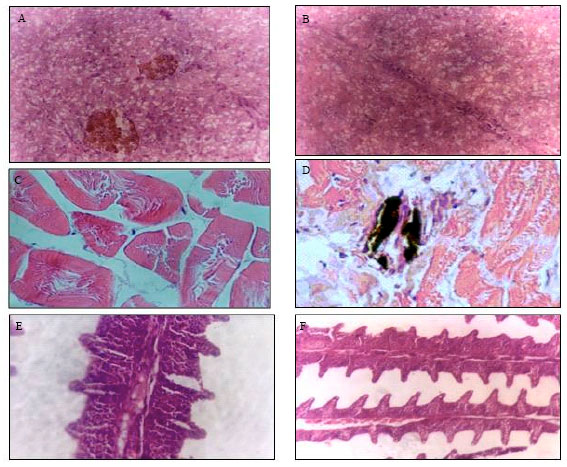 | |
| Fig. 2: | The histological changes in some vital organs exposed to 96 h LC50 concentration of hexavalent chromium (43.7 mg L‾1). A: Liver showing vacuolar degeneration with activation of melanomacrophage centers (H and E X400), B: Liver showing vacuolar degeneration with congestion and dilation of sinusoids (H and E X400), C: Histological section of muscle showing destruction and vaculation of the muscle cell (H and E X400), D: Histological section of muscle showing destruction and vaculation of the muscle cells with presence of melanomacrophages (melanophores) in the dermal layer (H and E X400), E: Gills showing congestion and hyperplasia of secondary lamellar cells with separation between the lamellar epithelial cells and capillary beds (H and E X400) and F: Gills showing severe congestion and hyperplasia of secondary lamellar cells especially at the base of the lamellae (H and E X400) |
Histological section of muscle in chromium-exposed fish showing destruction and vaculation of the muscle cell (Fig. 2C) and destruction and vaculation of the muscle cells with presence of melanomacrophages (melanophores) in the dermal layer (Fig. 2D)
Gills are the first target organ of several pollutants because of their very large interface area between external and internal fish environment, performing vital functions such as gas exchange and ion osmoregulation, the gills are particularly sensitive to adverse environmental conditions. In this study, gills were seen with congestion and hyperplasia of secondary lamellar cells with separation between the lamellar epithelial cells and capillary beds (Fig. 2E) and severe congestion and hyperplasia of secondary lamellar cells especially at the base of the lamellae (Fig. 2F).
REFERENCES
- Abbas, H.H., 2006. Acute toxicity of ammonia to common carp fingerlings (Cyprinus carpio) at different pH levels. Pak. J. Biol. Sci., 9: 2215-2221.
CrossRefDirect Link - Ambrose, T., A.K.L. Cyril, S. Vincen and L. Roselyn, 1994. Biochemical responses of Cyprinus carpio communis to the toxicity of tannery effluent. J. Ecobiol., 3: 213-216.
Direct Link - Bancroft, J., A. Stevens and D. Turner, 1996. Theory and Practice of Histological Techniques. 4th Edn., Churchill, Livingston, New York, London, San Francisco, Tokyo Pages: 740.
CrossRefDirect Link - Beyersmann, D., C. Block and A. Malviya, 1994. Effects of cadmium on nuclear protein kinase C. Environ. Health Perspect., 102: 177-180.
Direct Link - Blasiak, J. and J. Kowalik, 2000. A comparison of the in vitro genotoxicity of tri-and hexavalent chromium. Mutat. Res., 469: 135-145.
Direct Link - Claxton, L.D., V.S. Houk and T.J. Hugles, 1998. Genotoxicity of industrial wastes and effluents. Mutat. Res., 410: 237-243.
Direct Link - Dayan, A.D. and A.J. Paine, 2001. Mechanisms of chromium toxicity, carcinogenicity and allergenicity: Review of the literature from 1985 to 2000. Hum. Exp. Toxicol., 20: 439-451.
Direct Link - De Zwaan, A. and D.I. Zandee, 1972. Body distribution and seasonal changes in the glycogen content of the common sea mussel Mytilus edulis. Compa. Biochem. Physiol. Part A: Physiol., 43: 53-55.
CrossRefDirect Link - Gatta, P.P., A. Piva, M. Paolini, S. Testi, A. Bonaldo, A. Antelli and A. Mordenti, 2001. Effects of dietary organic chromium on gilthead seabream (Sparus aurata L.) performances and liver microsomal metabolism. Aquacult. Res., 32: 60-69.
Direct Link - Gomaa, M.N.E., A.A.K. Abou-Arab, A. Badawy and N. Khayria, 1995. Distribution pattern of some heavy metals in Egyptian fish organs. Food Chem., 53: 385-389.
CrossRef - Hartley, D.P., J.A. Ruth and D.R. Petersen, 1995. The hepatocellular metabolism of 4-hydroxynonenal by alcohol dehydrogenase, aldehyde dehydrogenase and glutathione S-transferase. Arch. Biochem. Biophys., 316: 197-205.
Direct Link - James, R., K. Sampath, K.V. Siva, S. Babu and P. Shanmuganandam, 1995. Toxic effects of copper and mercury on food intake, growth and proximate chemical composition in Heteropneustes fossilis. J. Environ. Biol., 16: 1-6.
Direct Link - Jiraungkoorskul, W., E.S. Upatham, M. Kruatrachue, S. Sahaphong, S. Vichasri-Grams and P. Pokethitiyook, 2003. Biochemical and histopathological effects of glyphosate herbicide on Nile tilapia (Oreochromis niloticus). Environ. Toxicol., 18: 260-267.
CrossRefDirect Link - Lemos, N.G., A.L. Dias, A.T. Silva-Souza and M.S. Mantovani, 2005. Evaluation of environmental waters using the comet assay in Tilapia rendalli. Environ. Toxicol. Pharmacol., 19: 197-201.
Direct Link - Majer, B.J., D. Tscherko, A. Paschke, R. Wennrich, M. Kundi, E. Kandler and S. Knasmuller, 2002. Effects of heavy metal contamination of soils on micronucleus induction in Tradescantia and on microbial enzyme activities: A comparative investigation. Mutat. Res., 515: 111-124.
CrossRef - Matsumoto, S.T. and M.A. Marin-Morales, 2004. Mutagenic potential of the water of a river that receives tannery effluent using the Allium cepa test system. Cytologia, 69: 399-408.
Direct Link - Matsumoto, S.T., M.S. Mantovani, M.I.A. Malaguttii, A.L. Dias, I.C. Fonseca and M.A. Marin-Morales, 2006. Genotoxicity and mutagenicity of water contaminated with tannery effluents, as evaluated by the micronucleus test and comet assay using the fish Oreochromis niloticus and chromosome aberrations in onion root-tips. Genet. Mol. Biol., 29: 1415-4757.
Direct Link - Mitchelmore, C.L. and J.K. Chipman, 1998. DNA strand breakage in aquatic organisms and the potential value of the comet assay in environmental monitoring. Mutat. Res./Fund. Mol. Mechan. Mutagen., 399: 135-147.
CrossRefDirect Link - Mount, D.R. and J.R. Hockett, 2000. Use of toxicity identification evaluation methods to characterize, identify and confirm hexavalent chromium toxicity in an industrial effluent. Water Res., 34: 1379-1385.
Direct Link - Negilski, D.S., 2002. Acute toxicity of zinc, cadmium and chromium to the marine fishes, yellow-eye mullet (Aldrichette forsteri C. and V.) and small-mouthed hardyhead (Atherinasoma microstoma Whitley) Aus. J. Mar. Freshwater Res., 27: 137-149.
CrossRefDirect Link - O'Brien, T., J. Xu and S.R. Patierno, 2001. Effects of glutathione on chromium-induced DNA crosslinking and DNA polymerase arrest. Mol. Cell Biochem., 222: 173-182.
Direct Link - Pan, Q., S. Liu, Y. Tan and Y. Bi, 2003. The effect of chromium picolinate on growth and carbohydrate utilization in tilapia, Oreochromis niloticus×Oreochromis aureus. Aquaculture, 225: 421-429.
CrossRefDirect Link - Passow, H., A. Rothestein and T.W. Clarkson, 1961. The general pharmacology of heavy metals. Pharmaocol. Rev., 13: 183-224.
PubMedDirect Link - Plotsky, Y., M.G. Kaiser and S.J. Lamont, 1995. Genetic characterization of highly inbred chickens line by two DNA methods: DNA fingerprinting and polymerase chain reaction using arbitrary primers. Anim. Genet., 26: 163-170.
Direct Link - Roling, J.A. and W.S. Baldwin, 2006. Alterations in Hepatic Gene Expression by Trivalent Chromium in Fundulus heteroclitus. Mar. Environ. Res., 62: S122-S127.
Direct Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Sastry, K.V. and K. Sunita, 1982. Toxic effects of chromium in a fresh water teleost fish, Channa punctatus. Punctatus. Toxicol. Lett., 11: 17-21.
CrossRef - Singh, J., L.C. Bridgewater and S.R. Patierno, 1998. Differential sensitivity of chromium-mediated DNA interstrand crosslinks and DNA-protein crosslinks to disruption by alkali and EDTA. Toxicol. Sci., 45: 72-76.
Direct Link - Sridevi, B. and S.L.N. Reddy, 2000. Effect of trivalent and hexavalent chromium on carbohydrate metabolism of a freshwater field crab, Barytelphusa guerini. Environ. Monitor. Assess., 61: 293-302.
Direct Link - Stohs, S.J. and D. Bagchi, 1995. Oxidative mechanisms in the toxicity of metal ions. Free Radic. Biol. Med., 18: 321-336.
CrossRefPubMedDirect Link - Sugiyama, M., 1992. Role of physiological antioxidants in chromium (VI)-induced cellular injury. Free Radical Biol. Med., 12: 397-407.
CrossRefPubMedDirect Link - Syversen, T.L.M., 1981. Effects of rnethyl mercury on protein synthesis in vitro. Actal. Pharmacol. Toxicol., 49: 422-426.
PubMedDirect Link - Von Burg, R. and D. Liu, 1993. Chromium and hexavalent chromium. J. Applied Toxicol., 13: 225-230.
CrossRefDirect Link - White, P.A. and J.B. Rasmussen, 1998. The genotoxic hazards of domestic wastes in surface waters. Mutat. Res., 410: 223-236.
Direct Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link