
Felicite Obono Mba
Department of Environmental Science, School of Resource and Environmental Science,
Wuhan University, Wuhan 430079, China
Xiong Zhi-Ting
Department of Environmental Science, School of Resource and Environmental Science,
Wuhan University, Wuhan 430079, China
Qiu Hai-Jie
Department of Environmental Science, School of Resource and Environmental Science,
Wuhan University, Wuhan 430079, China
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 18 | Page No.: 3065-3071







ABSTRACT
To test the effects of Salicylic Acid (SA) on physiological changes of plants under cadmium stress one cultivar of Chinese cabbage (Brassica chinensis) cadmium resistance, Changkenxiaobaicai (tolerance) were studied with pots cultures. Like other organisms Plants have adaptatives mechanisms whereby they are able to respond to both nutrient deficiency and toxicities. Plants materials were originated from the vegetable market in Wuhan, Hubei China. The seeds were tested on the pot cultures in the green house. The results of our experiments were quite different between the treatments. The combined treatment Cd 5 ppm + SA 500 mmol L-1 during our experiment showed that the activity of peroxidase and superoxidase dismutase in the cabbages were induced, the total chlorophyll content increased significally by 25.38%, chl a/b about 43.01% and total biomass about 41.67%, both in comparison with the control. The soluble sugar content increased significantly to about 25.47% in comparison with the control. The electrolyte leakages were less affected. Under SA treatment only, the chlorophyll content, chl a/b content increased to about 18.85%, in comparison with the control. Plant biomass increased about 18.90% with the addition of SA in the culture. SA treatment can increase or decrease the Chinese cabbages metabolism. Plants were exposed to 5 ppm CdCl2 for 5 days under natural light. Then they were cultivated with Hoagland nutrient solution, which served as control, nutrient solution supplemented with 5 ppm CdCl2. Five days after Cd treatment, seedlings were harvested. The experiment was performed in triplicate.
PDF Abstract XML References Citation
How to cite this article
Felicite Obono Mba, Xiong Zhi-Ting and Qiu Hai-Jie, 2007. Salicylic Acid Alleviates the Cadmium Toxicity in Chinese Cabbages (Brassica chinensis). Pakistan Journal of Biological Sciences, 10: 3065-3071.
DOI: 10.3923/pjbs.2007.3065.3071
URL: https://scialert.net/abstract/?doi=pjbs.2007.3065.3071
DOI: 10.3923/pjbs.2007.3065.3071
URL: https://scialert.net/abstract/?doi=pjbs.2007.3065.3071
NTRODUCTION
Cadmium (Cd) is easily transferred via the food chain. It is a highly toxic metal and has been ranked number 7 among the top 20 toxins mainly due to its negative influence on the enzymatic systems of cells (Al-Khedhairy et al., 2001; Sanita et al., 1999). Research has shown the toxic effect of cadmium on plants growth inhibition is a general phenomenon associated with cadmium (Collins et al., 1994). Cd interferes with many cellular functions mainly by complex formation with side group of organic compounds resulting in inhibition of essential activities. Although the mechanism of cytoplasm toxicity are identical in all organisms, different plants species and varieties show a wide range of plasticity in Cd tolerance, from the high degree of sensitivity of most plants on the one hand to the hyper accumulating phenotype of some tolerant higher plants on the other hand (Mc Grath et al., 2001). On an expanded concentration even sensitive species vary considerably in their response to Cd. pea for example, is considerably more sensitive to Cd than barley and barley is more resistant than Chinese cabbage which still grow well at concentrations above 10 μM Cd under nutrient solution.
Salicylic Acid (SA) is one kind of plant hormones, which has been widely used in agriculture and it has been reported that SA could also alleviate the cadmium toxicity on plants. In the early 1990s, it became apparent that SA is an endogenous compound that operates in the signaling pathway, for plant defense. Salicylic acid (SA) has been identified as an important signaling element involved in establishing the local and systematic disease resistance response of plants after pathogen attacks (Brune and Dietz, 1995; Alverez, 2000; Enyedi et al., 1992). After a pathogen attacks, SA levels often increase and induce the expression of pathogenesis related protein and initiate the delicate balance between functions.
For several years, it was known that, the application of SA or aspirin to tobacco induced gene expression and enhanced resistance to pathogens such as tobacco mosaic virus. SA plays a key role in plant disease resistance and hypersensitive cell death, but is also implicated in hardening response to abiotic stressors. SA accumulates to high levels at the site of infection with a subsequent but much smaller rise in the uninfected systemic tissues. The molecular events involved in SA signaling are not yet fully understood, although an increasing number of potential involved components, such as protein are being identified by molecular approaches (Klessing and Malamy, 1994). The early-proposed mode of SA action was related to the inhibition of Ascorbate peroxidase a major H2O2 scavenging enzyme. The inhibition might cause the cellular concentrations of H2O2 to rise subsequently; H2O2 may act as second messenger and activating defense-related genes (Shier, 1963). SA has broad divergent effects on stress accumulation and development damage of plants.
Therefore, the main objective of the present research is to study the influence of cadmium stress on the physiological changes in two populations of Brassica chinensis. Chinese cabbages and to analyze the possible reasons for their different tolerance to cadmium, This was addressed by measuring the total chlorophyll and biomass contents, peroxidase and superoxidase dismutase activity, electrolyte leakage. It is demonstrated that SA application caused partial protection against Cd toxicity in Brassica chinensis.
MATERIALS AND METHODS
Test soil and test plant species: Soil was collected in Wuhan university area, sieved and dried for 48 h at 150°C in the dryer machine. Soil ph was measured with a digital pH meter (model 420A; Oron research Inc., Boston, USA) after shaking 10 g of soil in 25 mL of pure water for 1-2 min and it let rested for half and hour and cover to avoid the contact with NH3 acid in air content. The total concentration of heavy metal in test soil was determined by acid digestion. Nitric acid (65%, 0.6 mL) and hydrochloric acid (35%, 1.8 mL) were added to 0.1 g of dry soil sample in a beaker and after which it was heated on the hot plate and cooled after addition of 0.1 M HCl and measured.
Plant material and preculture
Experiment 1: Chinese cabbage was utilized to carry out our experiments Brassica chinensis a common vegetable species were selected for experiments. Seeds were purchased from the local vegetable market in Wuhan China, sterilized in 3% HCHO for 5 min, washed with redistilled water for three changes and soaked in water over night. The pots were watered daily until seed germination. These were grown in twenty-four plastic pots for the Cd experiment with three replicates for each treatment. Four days later, seedlings were transferred to plastic pots containing 1000 g of soil. Plants were grown on half strength Hoagland’s solution containing macronutrients (mol m-3) 2.5 Ca (NO3)2, Ca (NO3)2. 4H2O, 5 KNO3, 1.0 KH2PO4, 2MgSO4, FeSO4.7H2O and micronutrients (mmol m-3) 1.0 Fe-EDTA. Thus control plants were supplied with water. Salicylic Acid (SA) was also sprayed every three days when the nutrient solution was replaced. The young seedlings were then grown under controlled cultural facilities at the temperature of 15-25°C and relative humidity of 70±3% in the grass house.
Complete nutrient experiment and cadmium treatments: When the seedlings were developed roots, one part was transported to the sand culture pots, after germination and appearance of 4 leaves, 4 seedlings were choose, washed and put in the pot containing soil (11 cm de diameter) contain 500 mL of nutrient solution and were covered with a foam plastic plate. Each plate was drilled with four evenly arranged, 5 mm diameter holes and one hole facing one plant. The roots were immersed in the solution. Adding 5 ppm Cadmium varied nutrient strength. Every 3 days, the culture solution were renewed distilled water were added to the pots at the same time with Cd solution of necessary, to compensate for the solution loss due the evaporation and transpiration. The plants were harvested one month after treatment, plant roots were washed with 5 mM L-1 Ca (NO3) 2 for 10 min to desorbs the Cd adsorbed on the root surface and further washed with distilled water.
Biomass and cadmium content measurement: The experiments were terminated after the plants had been exposed to metal treatments for 30 days.The plant samples were separated into leaves and roots. Fresh and dry weight of different parts of the plants was recorded. The base of roots and stems were rinsed thoroughly with double distilled water, then were put for 10 to 20 min in 5 mmol L-1 Ca (NO3)2 to desorbs the Cd absorbed on the roots surface and were washed one more time with distilled water blotted dry and then oven dry at 60°C for 48 h. Then biomass (DW) was determined. The dry plant tissues were cut into small pieces to mix them and digested in concentrated HNO3 first at room temperature for over night and then under a heating condition until the tissues particles dissolved. The samples were further digested with a mixture (HNO3: HClO4, 5:3v/v) and heat in an oven at 200-220°C. After cooling the extract were diluted and made up to 10 mL with 1 M HNO3 Cd concentration of the extract were determined Flame Atomic Absorption Spectrophotometer (FAAS) Polarized Zeeman (model Z-5000).
Electrolyte leakage: Fourteen days after Cd treatment, electrolyte leakage was estimated. Fresh leaf sample (0.1 g FW) was cut into a disk (1 cm diameter) and placed in a test tube containing 10 mL of deionized water. The tubes were then covered with plastic caps and placed in a water bath maintained at the constant temperature of 32°C for 2 h. Then the electrical conductivity of the solution (EC1) was measured using an electrical conductivity meter (model DDS-11A). The sample was then autoclaved at 120°C for 20 min and cooled, the electrical conductivity (EC2) was measured. The electrolyte leakage was expressed following the formula El = EC1/EC2x100
Chlorophyll determinations: Primary leaves were cut at their base from the 21 days old seedlings grown in soil culture, they were washed with fresh and later with double distilled water an amount of 0.5 g of the leaves were then put in the beaker, 10 mL of the solution (80% ethanol, 20% acetone) were added and the whole were let rest for 12-24 h and after that, the filter were done and 0.5 mL of the solution was mixed with 2.5 mL of (acetone +ethanol solution). Chlorophyll (a) and (b) were determined through the spectrophotometer (UV-9100) the content of chlorophyll (a) and (b) were estimated according to experimental equations:
Ca = 12.7A663 – 2.69A645 Cb = 22.9A645 – 4.68A663 Ca+b = 8.02A663+20.20A645 |
Antioxidant enzyme assay: Five days after Cd treatment, peroxidase (POD) activity was determined using the guaiacol oxidation method (Evans, 1965) for extraction of enzyme; leaf tissues (0.5 g Fresh Weight, FW) were homogenized with 0.1 M Tris-HCl buffer in a chilled pestle and mortar. The homogenate was centrifuged at 4000 rpm for 15 min and the resulting supernatant was used for determination of POD activity. POD activity was measured in a reaction mixture (4 mL) that contained 0.1 M phosphate buffer (pH 6.0) 1.6 μL H2O2, 1.5 μL guaiacol and 1 mL enzyme extract. The increase in absorbance was recorded at 470 nm in the UV-Vis light spectrophotometer (model UV-9100) within 30 sec (linear phase) after H2O2 was added, a unit of peroxidase activity was expressed as the change in absorbance per minute and per gram fresh weight of tissues.
Five days after Cd treatment, superoxidase dismutase (SOD) activity was measured according to Giannoplities and Ries (1965). Leaf tissues (0.5 g FW) were homogenized with 0.1 M sodium phosphate buffer (pH 7.8) in a chilled pestle and mortar. The homogenate was centrifuged at 4000 rpm for 20 min and the resulting supernatant was used for the determination of SOD activity. SOD activity was measured in a 4 mL reaction mixture that contained 0.1 M phosphate buffer (pH 7.8), 13 μM methionine, 7.5 μM nitro-blue tetrazolium, 2 μM riboflavin, 0.1 M EDTA (ethylene-diaminetetraacetic acid) and 1 mL enzyme extract. Absorbance by the reaction mixture was recorded at 560 nm in the UV-Vis light spectrophotometer (model UV-9100) and the activity was expressed as delta A560/(min g-1 FW).
Statistical analysis: The significance of differences between treatments was determined using t-test at p≤0.05 to test the effect of strength and Cd on plant biomass, roots length and Cd concentration in plant tissues. ANOVA (analysis of variance) was used to examine the separated effect of salicylic acid on roots length, plant biomass, sugar content, protein, chlorophyll and the Cd concentration in tissues. p≤0.05 means plant biomass and Cd content were compared with Least Significant Difference (LSD).
RESULTS
Plant biomass: Significant differences in plant biomass were observed between the treatments, in control without Cd and SA treatment, the total biomass is higher than the control (Table 1), however, when SA is sprayed individually in the cabbages leaves, the total biomass is higher and lower when Cd was adding alone, but when Cd is adding in association with SA in the culture, the total biomass increased.
Electrolyte leakage: The membrane damage is assessed by the conductivity of solute leakage from the cells. Electrolyte leakage from the leaves subjected to cadmium stress is shown (Fig. 1). Chinese cabbages treatments compared to each other shows that the cabbages leaves were more conductible with the addition of cadmium. When Cd +SA were added in the culture, the conductivity were significant (p = 0.004). The electrolyte leakage of the cabbages is correlated with the addition of Cd in the culture and its lower than the control.
Chlorophyll contents: Total Chlorophyll content in Brassica chinensis reduction is observed in all the treatments (p≤0.05). For Cd +SA treatments, the total chlorophyll content decreased about 25.38% in comparison with the control. When the seedlings were exposed to Cd only, the reduction was significant compared to the control (Fig. 2).
| Table 1: | Total Biomass content (mg plant-1 DW) in the cabbages (Brassica chinensis) after 3 months under culture in cadmium amended soil |
Results are means±SD (n = 3) (p<0.05) F1 = 6.922 | |
There is significant change of chlorophyll a/b content under Cd+SA treatment. When the cabbages were exposed to Cd and SA individually, chl a/b content was decreasing compare to the control. The chl a/b ratio decreased with Cd treatment that means chlorophyll (a) content is higher than chlorophyll (b) (Fig. 3). But under Cd + SA treatment, the chl a/b increased significally (p = 0.006), that means SA can reduce Cd stress by increasing the chlorophyll content.
Antioxidant enzyme: The POD activity increased significally with the addition of Cd in the solution of the Chinese cabbages p = 0.003. When SA is added in the culture, the enzyme activity decrease significantly the SA may alleviate Cd stress in the cabbages by reducing POD activity (Fig. 4).
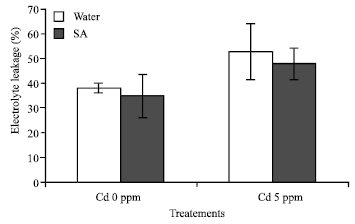 | |
| Fig. 1: | Electrolyte leakage of Brassica chinensis subjected 14 days of Cd treatment. An asterisk denotes that differences are statically significant (t-test p = 0.05); The results means±SD (n = 3); F2 = 5.413* cabbages populations p<0.03; F3 = 6.046*p<0.01; F1x3 = 3.404* p<0.05 |
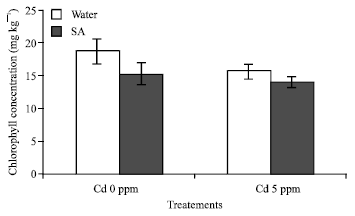 | |
| Fig. 2: | Chlorophyll content of Brassica chinensis subjected 21 days of Cd treatment. An asterisk denotes that differences are statically significant (t-test p = 0.05); The results means±SD; F2 = 157.722*** p<0.01; F3 = 4.668* p<0.05 |
The increase of SOD activity is observed with the addition of SA in the culture both for the Cd 0 and Cd 5 ppm (Fig. 5).
Element contents on roots and leaves: Cadmium content of leaves is higher under Cd + SA treatment than in the control in plant tissues (Fig. 6). The Cd concentration increased with the presence of Cd in the culture, but decrease with the addition of SA, Cd content in the leaves decrease while he is increasing in the roots. Cd adding individually in the cabbages increased but when SA was sprayed, the Cd content decreased. Differences in Cd content are observed between the treatments under SA treatment; the Cd content is higher than in the control. On the other hand Cd content in the root is significally higher than in the leaves this result indicate that, SA can reduce Cd content in the leaves and accumulation in the roots (Fig. 7).
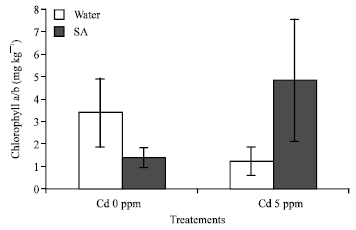 | |
| Fig. 3: | Chlorophyll a/b content of Brassica chinensis Results means±SD; F2 = 12.746*; p<0.05 |
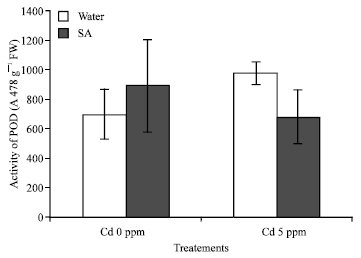 | |
| Fig. 4: | Activity of pod of B. chinensis results means (n = 3) ±SD average of three replicates; F2 = 7.072**; p<0.01 |
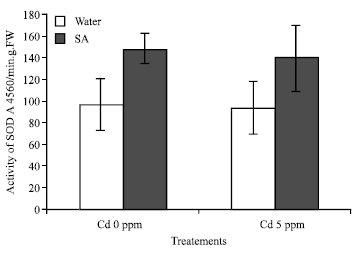 | |
| Fig. 5: | Activity of SOD of B. chinensis subjected 5 days after Cd treatment. An asterisk denotes that differences are statistically significant (t-test p<0.05; days results means±SD; F1 = 4.349*** chemical material; F2 = 8.011** cabbages populations; p<0.01 |
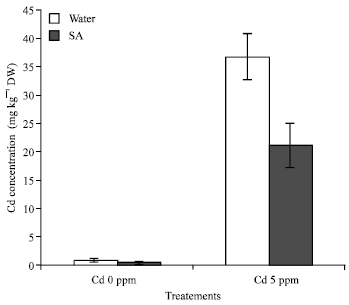 | |
| Fig. 6: | Cadmium concentration (mg kg-1 DW) in plant tissue of B. chinensis populations 30 days after culture in cadmium culture in amended soil. F3 = 24.90** p<0.01 |
SA can then tolerate the accumulation of Cd in the roots, this result is quite different from previous research which shows that leaves are an accumulator of heavy metals especially Cd. Its uptake increased with decreasing soil pH, Cd appears to be absorbed passively the difference in plant to accumulate heavy metals have been related to differences in their morphology.
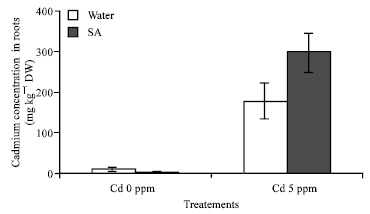 | |
| Fig.7: | Cadmium concentration (mg kg-1 DW) in plant tissue B. chinensis populations after 30 days of culture in cadmium culture in amended soil. F3 = 26.819** p<0.01 |
DISCUSSION
In general, plants posses physiological mechanisms that enable them to resist higher substrate heavy metal concentration (Chen et al., 1993). The current experiment as demonstrated that SA might play a key role in reducing Cd stress in Chinese cabbages. It is generally agreed that metal concentrations in plants vary with plant species (Cutler and Rains, 1974). Leafy vegetables tend to accumulate higher metal concentration than roots. The tolerance plant seems to possess stronger ability to exclude Cd from the roots to the leaves. Both roots and leaves absorbed metals and that translocation is low and availability of heavy metals in soil and their uptake by plants not only was dependent to the total metal content in the soil but also upon other factors (Antonovics et al., 1971; Maclan, 1976; Strickland et al., 1979; Adriano et al., 1986; Bjerre and Malamy, 1994). The differences in plant to accumulate heavy metal have been related to differences in their roots morphology (Miner et al., 1997). And our experiments confirm that metal concentration in leaves is much higher than in the roots, this heavy metal can be reduced in the leaves an accumulated in the roots. Cd content in the cabbages tissues increased in the addition of SA. In the current experiment, SA introduced in the culture may reduce the transport of Cd from roots to leaves (Fig. 6 and 7). Heavy metal and Cd generally affect water plant in particular where critically reviewed. Cd generally decreases the water stress tolerance of plants, causing turgor less at higher relative water content and leaf water potential than in on non-treated control plants. Cd increases the bulk elastic modulus and therefore decrease cell wall elasticity seems to be an important cause of the low water stress tolerance in the affected plants (Wierbicka, 1998).
Biomass alteration is regarded as a common response of plants to various stresses. The reduction of biomass at the Cd 5 ppm treatment is significally different (p≤0.05). The roots and leaves total biomass is increasing more evidently (Table 1), this indicated that, the pollutant concentration affected the water relation in the Chinese cabbages and can be correlated to the chlorophyll content difference (Fig. 2). This result is in accordance with the fact that, the capacity of absorption and transportation of water and nutrient had been inhibited due to Cd and SA stress in the cabbages.
The increasing of the electrolyte leakage in this study can assess the membrane damage; it’s observed that, the increasing electrolyte leakage in the leaves is significant; the higher level of electrolyte leakage is in correlation with the presence of Cd and SA simultaneously in the culture (Fig. 1). There is no direct correlation between the cabbages species and the Cd content in the leaves. The lower or the higher Cd content in tissues would be expected to induce stronger peroxidation of the membrane thus leaking to a greater electrolyte leakage. This explanation is supported by the increasing or decreasing POD and SOD activity in the cabbages. The POD activity in the cabbage can be also related to the Cd content in the leaves.
Decrease of chlorophyll content in the cabbages after exposure to cadmium is frequently reported in the literature (Hemphill et al., 1972; Miller et al., 1976). It is evident from the present study that Cd treatment result in a significant loss of chlorophyll (Fig. 2) present results also show that the total chlorophyll content in the cabbages is lower. These results might be related to the Cd content in the roots and this can cause sever damage of nutrient transportation from roots to leaves. The lower chlorophyll content would then result in lower biomass production; similar results were reported in Brassica laevigata (Chugh and Sawhney, 1999) and it as also demonstrated that, plant that are resistant to heavy metal generally have lower growth in comparison with the plant growth under heavy metal contaminated area (Hui et al., 2000). Cd toxicity in leaves caused a decrease in chlorophyll content but former works also shows that increasing concentration of Cd from 0.1-5 mM progressively decrease chlorophyll content (Wierbicka, 1998) the photosynthesis activity might be more higher and normal, chl a/b content is increasing in the cabbages specially under Cd and SA stress. We can also infer that the chlorophyll (a) is higher than the chlorophyll (b) (Fig. 3).
POD induction is a general response of higher plants to resist under heavy metal stress, his activity in the cabbages at Cd 5 ppm treatment is higher (Fig. 4), this could be the reason for the cabbages resist to Cd when SA is adding in the culture. The POD activity is lower but this could be also explained by stating that, SA can reduce Cd stress in the tolerance plant. This result is similar to other research results which conclude that, in some tolerance plants, the activity of POD is higher to enable the plant to protect themselves against the oxidative stress (Hsu and Koa, 2004) while as such high activities were not observed in non-tolerance plants (Prasad et al., 1995; Scalet et al., 1995), another reason for the low POD activity might be that the population accumulated a higher Cd content in leaves and this process could inhibit the POD activity.
Contrary to the POD, the activity of SOD significantly increased in the cabbages (Fig. 5) however at Cd 5 ppm, the activity of SOD in the cabbages can be also related to the Cd content in the leaves. The increasing of Cd in the leaves could exerted a positive effect in the SOD activity similar result were found in rice in which the resistance variety has higher SOD activity and lower metal content in leaves (Kronfuss et al., 1996), the addition of SA in the culture reduced significantly the SOD activity under Cd stress. The complex Cd + SA can also play a key role in the reduction of SOD activity in the cabbages. We have also realized that while the Cd concentration in the leaves is higher, the SOD activity is also higher and we can mention that the SOD activity is growing or decreasing with the Cd concentration in the leaves membrane damage caused by lipid peroxidation under Cd stress is mediated by activated oxygen radicals, like H2O2, hydroxyl and superoxide radicals, but these reactive molecules can be quenched by the induction of specific enzymes, like POD, SOD (Yamasaki et al., 1997).
According to all the above results, Cd can promote generation of some harmful free radicals, such as Reactive Oxygen Species (ROS) and peroxides compounds, which in turn can damage cell membrane (De Vos and Schat, 1981; Maribel and Satoshi, 1998). SA can alleviate some harmful stress in the cabbages such as reducing biomass, chlorophyll content, SOD and POD activities. Further studies have to be carried out in order to understand the particular effect of Cd and SA on the Chinese cabbages. From those results and according to the previous research, we can conclude that SA can be used to alleviate the cadmium toxicity and consequently improve the main nutritive quality of Chinese cabbages under cadmium stress.
REFERENCES
- Al-Khedhairy, A.A., S.A. Al-Rokayan and F.A. Al-Misned, 2001. Cadmium toxicity and cell stress response. Pak. J. Biol. Sci., 4: 1046-1049.
CrossRefDirect Link - Alverez, M.E., 2000. Salicylic acid in the machinery of hypersensitive cell death and disease resistance. Plant Mol. Biol., 44: 429-442.
PubMedDirect Link - Antonovies, J., A.D. Bradhava and R.G. Turner, 1971. Heavy metal tolerance in plants. Adv. Ecol. Res., 7: 1-85.
Direct Link - Brune, A. and K.J. Dietz, 1995. A comparative analysis of element composition of barley roots and leaves under cadmium-, molybdenum-, nickel-and zinc-stress. J. Plant Nutr., 18: 853-868.
Direct Link - Chen, Z., J.W. Riigliano and D.F. Klessig, 1993. Purification and characterization of a soluble salicylic acid-binding protein from tobacco. PNAS., 90: 9533-9537.
PubMedDirect Link - Chugh, K.L. and K.S. Sawhney, 1999. Photosynthetic activities of Pisum sativum seedlings grown in presence of cadmium. Plant Physiol. Biochem., 37: 297-303.
CrossRefDirect Link - Klessig, D.F. and J. Malamy, 1994. The salicylic acid signal in plants. Plant Mol. Biol., 26: 1439-1458.
CrossRefDirect Link - Dionisio-Sese, M.L. and S. Tobita, 1998. Antioxidant responses of rice seedlings to salinity stress. Plant Sci., 135: 1-9.
CrossRefDirect Link - Miner, G.S., R. Gutierrez and L.D. King, 1997. Soil factors affecting plants concentration of Cd, copper and zinc on sludge amended soils. J. Environ. Qual., 26: 989-994.
Direct Link - Prasad, M.N.V., 1995. Cadmium toxicity and tolerance in vascular plants. Environ. Exp. Bot., 35: 525-545.
CrossRefDirect Link - Scalet, M., R. Federice, M.C. Guido and F. Manes, 1995. Peroxidase activity and polyamine changes in response to ozone and simulated acid rain in Aleppo pine needles. Environ. Exp. Bot., 35: 417-425.
Direct Link - Wierbicka, M., 1998. The adaptation of Silene vulgaris to growth on a calamine waste heap (S. poland). Environ. Pollut., 101: 415-426.
CrossRefDirect Link - Yamasaki, H., N. Sakihama and N. Ikehara, 1997. Flavonoid-peroxidase reaction as a detoxification mechanism of plant cells against H2O2. Plant Physiol., 115: 1405-1412.
PubMedDirect Link