
Etou Mongo
Laboratoire de Biophysicochimie et Technologie Alimentaires. Ecole Nationale Superieure Polytechnique, Universite Marien Ngouabi, B.P. 69, Brazzaville-Congo
Antoine
Laboratoire de Biophysicochimie et Technologie Alimentaires. Ecole Nationale Superieure Polytechnique, Universite Marien Ngouabi, B.P. 69, Brazzaville-Congo
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 16 | Page No.: 2670-2675







ABSTRACT
Protein from flour and doughs prepared at high temperature in presence and absence of oxidants (potassium bromate, potassium iodate and L-ascorbic acid) was fractionated according to solubility into water, salt, alcohol, acetic acid, soluble protein fractions and insoluble residue protein. All fractions were freeze-dried and subjected to scanning electron microscopy to observe visually the changes in protein structure. Acetic acid-soluble and insoluble residue protein are alike in structure, but the former was thermally denatured easily, while the latter was very stable to heat treatment. Salt and alcohol, soluble protein were not deformed, but the water soluble protein was deformed by heat treatment in the absence of oxidant. Oxidants generally promoted deformation of protein structure with the exception that bromate partly protected acetic acid-soluble protein from deformation.
PDF Abstract XML References Citation
How to cite this article
Etou Mongo and Antoine, 2007. Study by Scanning Electron Microscopy of Mixture of Cereal Proteins Fractions (Maize and Wheat) from Bread Doughs Prepared at High Temperature in the Presence of Oxidants. Pakistan Journal of Biological Sciences, 10: 2670-2675.
DOI: 10.3923/pjbs.2007.2670.2675
URL: https://scialert.net/abstract/?doi=pjbs.2007.2670.2675
DOI: 10.3923/pjbs.2007.2670.2675
URL: https://scialert.net/abstract/?doi=pjbs.2007.2670.2675
INTRODUCTION
Cereal (maize and wheat) dough mixing is the most critical step in the breadmaking process. Many physicochemical changes in dough component combine to produce the overall effect of mixing; the flour proteins undergo the most important of these changes. Meckam et al. (1965) found that the amount of protein extracted from dough by dilute acetic acid increased with extended mixing. They postulated that the mixing decreased the size of protein aggregates in the flour particles. This mechanism was later supported by Tsen (1969). In contrast, Tanaka and Bushuk (1973) concluded, after separating more glutenin from dough than flour, that mixing resulted in polymerisatin of large insoluble proteins.
Because of the important role of wheat protein in the breadmaking quality of bread wheat, its microscopy structure might shown a recognizable relationship to its ability to contribute to baking quality. Rosell et al. (2003) reported that wheat flour proteins were affected by transglutaminase and glucose oxidase.
Seckinger and Wolf (1970). reported that gladian showed particle structures, whereas glutenin formed fibillar extensions. Orth et al. (1973) observed a fibrous and ribbon-like structure for purified glutenin from a few varieties of wheat. Chen et al. (1996) studied the effects of dough mixing and oxidizing improvers on free reduced and free oxidized glutathione and protein-glutathione mixed disulfides of wheat flour. Demiralp et al. (2000) studied on electrophoretic patterns of flour proteins during dough mixing, the effect of oxidizing agents and defatting. Aja Silvia et al. (2003) observed the improvement of cereal protein net works through enzymes. Bernadin and Kasarda (1973) reported that endosperm protein was in sheet form when wetted resulting from laminar disposition of the protein. Khoo et al. (1975) reported that the protein integrated to yield a continuous film-like matrix by dough mixing. Bechtel et al. (1978) observed that excessive mixing destroyed the matrix. Rasiah et al. (2004) noticed the cross linking of wheat dough proteins by glucose oxidase and the resulting effects on bread and croissants.
Recently, Tu and Tsen (1978) further clarified that glutenin stretched and spread to form a sheet-like structure with mixing and that the structure was reduced in size with prolonged mixing. In a previous study, the Do-Corder was employed to investigate the effect of potassium bromate, potassium iodate and L-ascorbic acid on dough mixed at high temperature (Tanaka et al., 1980). The Co-Corder curve from bromated dough showed two distinct peaks at 75 and 80°C, while that for the remainder had only one peak at either 85 or 75°C which were assigned to the consistency effects of gluten and starch, respectively.
This study was designed as a visual examination of the structural changes produced in dough protein by mixing with the Do-Corder in the presence and absence of the oxidants at elevated temperature at 75 and 85°C where distinct changes occurred in Do-Corder curves.
MATERIALS AND METHODS
Materials: This study was conducted at the Laboratory of Biophysico-chemical and Technology Alimentary of Compiegne University of Technology (France) in 2006. White maize was growed at Intsiala in district of Gamboma (Congo-Brazzaville). The maize flour was prepared at Brazzaville in the Congo Society of Cereals and red wheat flour prepared at french Society Mon Moulin. The bread flour was composed of 30% of white maize flour and 70% of red wheat flour. The flour mixed contained 12.1% of protein; 14.7% of moisture and 0.43% of ash. The oxidants used are: potassium bromate, potassium iodate and L-ascorbic acid.
Methods: All analysis of this study were carried out at the Laboratory of Biophysico-chemical and Technology Alimentary of Compiegne University Technology (France).
Preparation of dried doughs: Doughs containing 0 or 1200 parts 10-6 of oxidant were mixed in the Do-Corder and heated at a rate of about 3°C min-1 from 35°C until the appropriate temperature was reached (Tanaka et al., 1980). During this time, the dough consistency was recorded to produce the tracings. When a dough sample was to be taken, mixing and heating were stopped at the appropriate stage and the sample was quickly frozen by immersion in liquid nitrogen. The frozen dough was freeze-dried, pulverised by hand and ground in a coffee grinder to pass through an 80-mesh sieve.
Determination of sulfhydryl contents: Sulfhydryl (SH) contents of the flour and ground doughs were determined by an amperometric titration method developed by Sokol et al. (1959) and modified by Tsen and Anderson (1963). SH contents are expressed in μEq g-1 of dry sample.
Protein solubility: One gram of sample was placed into a 100 mL beaker and dispersed completely in 25 mL of 0.1 M acid acetic by stirring. The suspension diluted to 100 mL with 0.1 M acid acetic was kept overnight at room temperature before being centrifiged at 3000 g for 10 min. The soluble nitrogen was determined by the micro-kjeldhal method. The protein solubility was calculated as:
Extraction and fractionation of proteins: The proteins of the flour and ground dough samples were extracted and fractionated according to solubility by a modified Osborne procedure as described by Chen and Bushuk (1970). All the soluble fraction was further purified by dissolving in 0.05 M acetic acid. Occluded starch was removed by centrifugation at 25, 000 g for 25 min. The supernatant solutions were dialysed against 0.05 M acetic acid and freeze-dried. The insoluble residue protein was extracted from fraction with 0.01 M acetic acid, 3 M urea and 0.01 M cetyltrimethylammonium bromide by stirring overnight in a cold room at 4°C. This suspension was centrifuged for 25 min at 20, 000 g, the supernatant was dialysed against water and the precipitated protein freeze-dried. All the purified fractions were subjected to scanning electron microscopy.
Scanning electron microscopy: The dried samples were sprinkled on to double-sided adhesive tape attached to specimen stubs. The mounted specimens were coated with a layer of platinum about 15.0 nm thick in Eiko ioncoater of type IB-3 and viewed with a Hitachi S450 at 20 kV.
RESULTS
Sulfhydryl oxidation and protein denaturation: Denatured protein was defined as protein with decreased protein solubility. The SH content and protein solubility of the control and oxidised doughs decreased as the temperature was raised to 75°C (Table 1). This trend was extern for the SH content of bromated dough and no SH was detected. There was no further decrease in SH content for any sample with increasing temperature to 85°C. Protein solubility was slightly higher for the oxidised dough especially the bromated one than for the control at 85°C. Protein of the oxidised dough become more tolerant to heat than that of the control, presumably due to oxidation of SH to SS bonds to form a heat-stable conformation in the gluten. However, it is also possible that the added oxidants may have oxidised the SH groups to sulphinic, sulphonic or sulphenic acids.
Scanning electron microscopy: The scanning electron microscopy was employed to observe the effect of oxidants on the extent of change in structure for each protein fraction.
| Table 1: | The effect of oxidants on SH oxidation and protein denaturation in heated dough |
 | |
Water-soluble fraction: Scanning electron micrographs of water-soluble protein (albumin) are shown in Fig. 1. Albumin showed a needle or strand structure (Fig. 1a). This structure transformed to a sheet-form with increasing temperature (Fig. 1b). This change in the structure with heat treatment is probably caused by thermal denaturation of the protein. The change in the structure was slightly affected by the addition of oxidants (Fig. 1c). A sheet-form was in part transformed into a net-form may be due in breakdown.
Salt-soluble fraction: Scanning electron micrographs of salt-soluble protein (globulin) are shown in Fig. 2. Globulin had a droplet shape like starch with a diameter of 1-5 μm (Fig. 2a). Some needle-shaped contaminants can be seen here due to incomplete separation. This droplet shape was deformed by heat only in the presence of oxidant. Individual droplets are barely visible for the protein form bromated dough (compare Fig. 2b and c).
Alcohol-soluble fraction: Figure 3a shows a scanning electron micrograph of alcohol-soluble protein (gliadin)
extracted from flour. Gliadin appeared as a needle with a round tip, but also had a partly sheet-like structure. This structure transformed into a film only on the addition of oxidant specially iodate with increasing temperature (compare Fig. 3b and c).
Acid acetic-soluble fraction: Figure 4a shows a scanning electron micrograph of acetic acid-soluble protein (glutenin) extracted from flour. Glutenin possessed a network structure composed of narrow sheets and strands (fibrous) and was severely altered by heat treatment (Fig. 4d). The protein from the iodated and the ascorbated dough (Fig. 4d) treated at 85°C, also suffered extensive cleavage caused by thermal denaturation (compare Fig. 4b and c).
Residue fraction: The protein of the residual fraction possessed a structure similar to that of the acetic-soluble fraction (glutenin) in Figure 5a. There was no effect of heat treatment on the structure of residue protein was evident in the control dough (compare Fig. 5a and b). Oxidants, however, did affect the structure of the residue protein transforming it into a sheet or film-like structure (Fig. 5c).
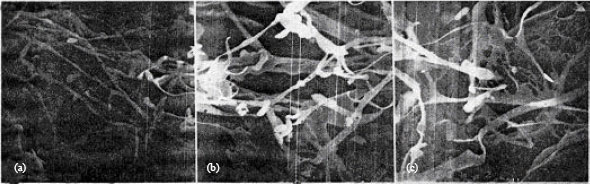 | |
| Fig. 1: | Scanning electron micrographs of water-soluble protein (albumin) from: (a) flour, (b) control dough at 75°C and (c) oxidants dough at 85°C |
 | |
| Fig. 2: | Scanning electron micrographs of salt-soluble protein (globulin) from: (a) flour, (b) control dough at 85°C and (c) bromated dough at 85°C |
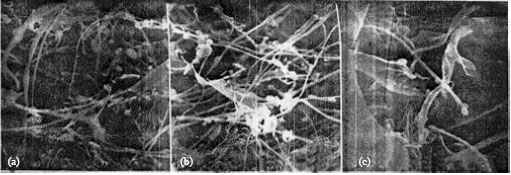 | |
| Fig. 3: | Scanning electron micrographs of alcohol-soluble protein (gliadin) from: (a) flour, (b) control dough at 85°C and (c) iodated dough at 85°C |
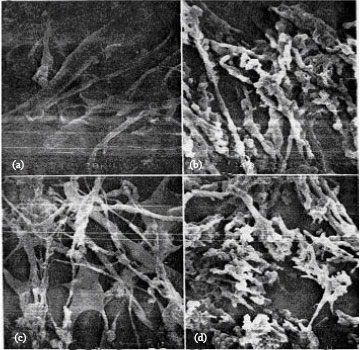 | |
| Fig. 4: | Scanning electron micrographs of acetic acid-soluble protein (glutenin) from: (a) flour, (b) control dough at 75°C, (c) bromated dough at 75°C and (d) ascorbated dought at 58°C |
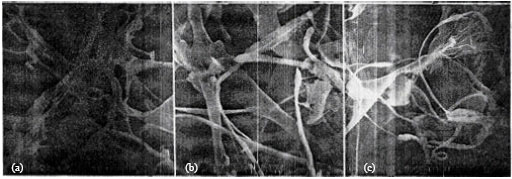 | |
| Fig. 5: | Scanning electron micrographs of residues protein from: (a) flour, (b) control dough at 85°C and (c) bromated dough at 85°C |
DISCUSSION
Seckinger and Wolf (1970) observed that on the spreading of wheat proteins on liquid surfaces gliadin dispersed into particles while glutenin remained together with fibrillar extensions. In the present study, however, freeze-dried gliadin and glutenin from a flour appeared needle-sharped and narrow sheet-or strand-shaped respectively. The observation for glutenin is in accordance with the results reported by Orth et al. (1973). The diversity among workers may be due to the difference in variety of wheat and in the method of glutenin preparation. According to the observation, oxidants promoted thermally induced changes in the structure of water-and salt-soluble protein. It was not clear in the study whether or not their oxidizing action on the water-and salt-soluble protein may play a role in breadmaking performance. Although Rasiah et al. (2004) reported that these two protein groups are not involved in baking performance. Alcohol-soluble protein was also transformed into a film-like shape in the presence of oxidants, specially iodate with increasing temperature.
Glutenin and residual protein are alike in structure (Fig. 4a and 5a). This is in accordance with the results of Orth et al. (1973) who concluded that the protein of the residual fraction is mainly high molecular weight glutenin. Both proteins however, behaved in quite different ways under heat treatment. Glutenin was thermally denatured easily but the residual protein was very stable to heat treatment. This might be positively related to the fact that residual protein plays an important role in the baking performance of a flour (Rosell et al., 2003; Orth and Bushuk, 1972). In the present study, oxidants generally promoted the deformation of protein structure with the exception that bromate partly protected glutenin from the deformation. For albumin, gliadin and residue protein from oxidisel doughs, the structure transformed into a like-film shape even when mixed at 85°C. This contrasts somewhat with the results of Tu and Tsen (1978) who reported that a film-like structure of aggregated glutenin, formed by an initial mixing was reduced in size with prolonged mixing. The film formation of proteins especially residue protein, by oxidants might play a critical part in improving the breadmaking quality of a flour.
CONCLUSIONS
The scanning electron microscopy was employed to observe with effectiveness the effect of oxidants in the modification of the protein structure in presence of high temperature. The present research provide clear evidence that oxidants added at a level 1200 parts 10-6 have a marked effect on each protein fraction in dough. The protein of the residual fraction possessed a structure similar to that of the acetic-soluble fraction (glutenin). Glutenin possessed a network structure composed of narrow sheets and strands and was severely altered by heat treatment. Gliadin appeared as a needle with a round tip, but also had a partly sheet-like structure. Globulin had a droplet shape like starch with a diameter of 1-5 μm.
REFERENCES
- Demiralp, H., S. Celik and H. Koksel, 2000. Effects of oxidizing agents and defatting on the electrophoretic patterns of flour proteins during dough mixing. Eur. Food Res. Technol., 211: 322-325.
CrossRefDirect Link - Rosell, C.M., J. Wang, S. Aja, S. Bean and G. Lookhart, 2003. Wheat flour proteins as affected by transglutaminase and glucose oxidase. Cereal Chem., 80: 52-55.
CrossRef