
Gholamali Akbari
Abourayhan Campus, University of Tehran, Tehran, Iran
Seyed Ali Mohammad Modarres Sanavy
Department of Agronomy, Faculty of Agricultural, Tarbiat Modarres University, P.O. Box 14115-336, Tehran, Iran
Saeed Yousefzadeh
Department of Agronomy, Faculty of Agricultural, Tarbiat Modarres University, P.O. Box 14115-336, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 15 | Page No.: 2557-2561







ABSTRACT
Seeds of three wheat (Triticum aestivum L.) cultivars included Mahdavi, Pishtaz and Shiraz, were used to investigate the effects of different salinity and auxin concentrations on their germination percent, radicle and hypocotyl length, seedling fresh and dry weight, radicle and hypocotyl dry weight. Results showed that increasing concentrations of NaCl reduced germination percentage, radicle length, hypocotyl length, seedling fresh and dry weight and hypocotyl dry weight. Hypocotyl dry weight increased in seeds only at -0.6 MPa. Auxin increased hypocotyl length, seedling fresh and dry weight and hypocotyl dry weight, but did not influence on seed germination percentage and radicle length. Pishtaz cultivar showed high seed germination percentage, radicle length and hypocotyl length in comparison to other cultivars while Shiraz cultivar had high radicle dry weights and Mahdavi cultivar produced high seedling fresh and dry weights.
PDF Abstract XML References Citation
How to cite this article
Gholamali Akbari, Seyed Ali Mohammad Modarres Sanavy and Saeed Yousefzadeh, 2007. Effect of Auxin and Salt Stress (NaCl) on Seed Germination of Wheat Cultivars (Triticum aestivum L.). Pakistan Journal of Biological Sciences, 10: 2557-2561.
DOI: 10.3923/pjbs.2007.2557.2561
URL: https://scialert.net/abstract/?doi=pjbs.2007.2557.2561
DOI: 10.3923/pjbs.2007.2557.2561
URL: https://scialert.net/abstract/?doi=pjbs.2007.2557.2561
INTRODUCTION
Seed germination is a crucial stage in the life history of plants and salt tolerance during germination is critical for the establishment of plants that grow in saline soils (Khan et al., 2000). Several investigations of seed germination under salinity stress have indicated that seeds of most species attain their maximum germination in distilled water and are very sensitive to elevated salinity at the germination and seedling phases of development (Gulzar et al., 2003; Ghoulam and Fares, 2001).
Salinity can affect growth, dry matter accumulation and yield (Sultana et al., 1999). It is well known that dry mass of plants is reduced in proportion to the increase in salinity (Romero-Aranda et al., 2001). Selecting cultivars for rapid and uniform germination under saline conditions can contribute towards early seedling establishment. Seeds of most halophytes attain their maximum germination in the absence of NaCl and are very sensitive to elevated salinity at the germination and early establishment phases (Khan et al., 2000).
Tolerance to salinity is not a fixed characteristic in plants and may vary with the stage of growth even within a species. In field crops that are relatively halo-tolerant, such as varieties of wheat (Triticum aestivum L.), seed germination is a major factor limiting success (Almansouri et al., 2001).
Numerous studies show that application of several plant growth regulators such as auxins may increase the germination ability of seeds and seedling vigour in a wide range of plants (Swaminathaan and Srinivasan, 1996; Balestri and Bertini, 2003). Information on the effects of auxins on seed germination is still limited, but there is evidence that exposure of seedlings of different plants to auxins may influence root development (Reed et al., 1998; Balestri and Bertini, 2003).
The presence of some auxins such as IAA in seed germination conditions, increase germination percentage and coleoptile elongation (Rekoslavskayal et al., 1999). The effect of different auxins on seed germination process include direct and indirect evidences indicating the involvement of auxins in the seed germination (Chiwocha et al., 2005).
Numerous studies have demonstrated improvement in seed germination of different plant species under both normal and stress conditions in response to priming with plant growth hormones or other organic substances (Ashraf and Foolad, 2005). In wheat, while seed germination decreased with increasing levels of salinity, the adverse effect of salinity was alleviated by soaking seed with IAA or NAA (Balki and Padole, 1982). In a different study with wheat, adverse effects of salinity stress on seed germination were mitigated by soaking seed in varying concentrations of IAA (Gulnaz et al., 1999). Salinity is one of the major limiting factors in crop production in arid and semiarid regions in many countries in the world like Iran. It is necessary to understand the response of wheat seeds to salinity stress if cultivation in saline areas is considered. Auxin is a plant hormone that can positive effect on germination percentage in crop seeds especially in salinity stress condition. Thus the aim of this study was to investigate the response of three wheat cultivars to different auxin and salinity levels during seed germination.
MATERIALS AND METHODS
Seeds of three cultivars of wheat, namely: Mahdavi, Shiraz and Pishtaz were used in this study. The trial was conducted at Faculty of Agriculture, Tarbiat Modarres University, Tehran, Iran, in 2005. Seeds were germinated in covered, sterilized, disposable petri dishes containing Whatman filter paper moistened with either distilled water (control), or different treatment solutions. Germination was assessed using three replicates of 50 seeds in a factorial laid out in Completely Randomized Design (CRD) testing combinations of three levels of salinity (0, -0.6 and -1.2 MPa NaCl, Table 1) (Braccini et al., 1996) and three levels of auxin (0, 1 and 2 mg LG1 IAA) on seeds of three wheat cultivars in 9 cm diameter Petri dishes.
Seeds were incubated in a growth chamber at 25°C and were considered germinated with the emergence of the radicle. Germination was scored when a 2 mm radicle had emerged from the seed coat. Every three days, the germinated seeds were removed from the Petri dishes. The seeds to germinate in each replicate were retained for measurements of radicle and hypocotyl lengths at the end of the experiment. After 240 h, final germination percentages were recorded and seedling fresh weights immediately determined. To determine the impact of the treatments on seed germination, all seedlings were separated from the remaining seeds. Seedling dry weights were evaluated after 48 h in an oven at 80°C.
In order to maintain adequate moisture, 5 mL of the original salt solutions were added to each Petri dish every three days. Germination percentage, radicle and hypocotyl lengths, seedling fresh and dry weights, radicle and hypocotyl dry weights and total dry weight were measured. Data were analyzed using analysis of variance (ANOVA) and Duncan=s multiple range test (for comparison the treatment means) and using SAS software version 6.12 (SAS Institute Inc. 1996).
| Table 1: | Used amounts of sodium chloride to obtain different levels of water deficit |
 | |
RESULTS AND DISCUSSION
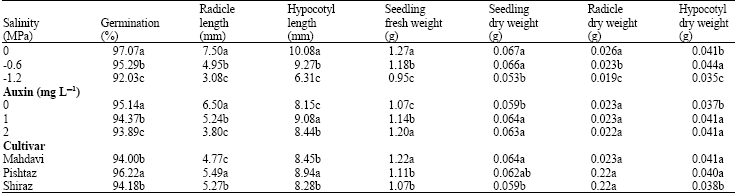
Analysis of variance revealed significant differences among wheat cultivars, salinity and auxin levels for germination percentage, radicle and hypocotyl lengths (p<0.01), seedling fresh and dry weights (p<0.01) and hypocotyl dry weight (p<0.01) (Table 1). Salinity affected on radicle dry weight (p<0.01) but hormone had not significant effect on this trait. The interaction effects of SalinityHCultivar (SHC), HormoneHCultivar (HHS), Salinity HHormone (SHH) and SalinityHHormoneHCultivar (SHHHC) were significant for all traits except hypocotyl dry weight. Also interaction effect of SHC was not significant for seedling dry weight.
Increasing concentrations of NaCl reduced germination percentage, radicle length, hypocotyl length, seedling fresh and dry weights, radicle and hypocotyl dry weight, but increased hypocotyl dry weight at the potential -0.6 MPa (Table 2).
Auxin enhancement increased hypocotyl length, seedling fresh and dry weights and hypocotyl dry weight but, reduced seed germination percentage at 0.6 salinity level in comparison with control (Table 2). These results are consistent with Rekoslavskayal et al. (1999), where, application of some auxins such as IAA in seed germination increased coleoptile elongation. Thus, seed priming with auxin had positive effects on some growth traits of wheat seeds. There is also evidence of growth under saline conditions by using plant growth hormones as priming agents, such as GA and ascorbic acid in wheat (Al-Hakimi and Hamada, 2001). Auxin concentrations were not significant on radicle dry weight (Table 2).
Pishtaz cultivar showed high seed germination percentage, radicle and hypocotyl length traits in comparison to other cultivars while Mahdavi cultivar produced high seedling fresh and dry weights (Table 2). There were not significant differences among wheat cultivars for radicle dry weights.
Germination percentage reduced at the highest level of salt (-1.2 MPa). At 2 mg LG1 auxin with no salt, the Pishtaz cultivar germination percentage was higher than other tested cultivars (Table 3). These results were in agreement with Xue et al. (2004) who found that high levels of salinity can significantly inhibit seed germination. Further, Waisel (1972) found that increasing salinity concentrations in germination often cause osmotic and/or specific toxicity which may reduce or retard germination percentage. Salt induced inhibition of seed germination could be attributed to osmotic stress or to specific ion toxicity (Huang and Redmann, 1995).
| Table 2: | Means values (main effects) of seed germination traits for three wheat cultivars auxin and salinity stress |
 | |
| Means with similar letter (s) in each trait is not significantly different at 5% probability level according to Duncan's Multiple Range Test | |
It has been reported by several authors that salinity stress affects seed germination either by decreasing the rate of water uptake (osmotic effect) and or facilitating the intake of ions, which may change certain enzymatic or hormonal activities inside the seed (ion toxicity) (Huang and Redmann, l995).
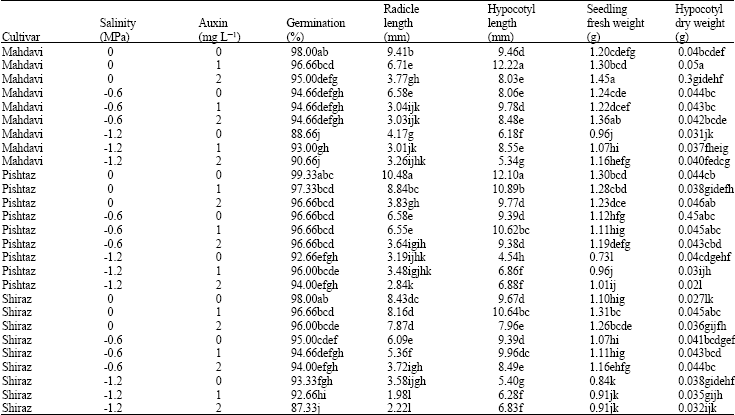
Seed germination percentage was increased in high salinity conditions with presence of 1 mg LG1 auxin for Mahdavi and Pishtaz cultivars. Also presence of 1 mg LG1 auxin for Shiraz cultivar was not decreased seed germination percentage in higher level (-1.2 MPa) of salinity (Table 3). Wheat plants grown under saline conditions from seed soaked in IAA, NAA and GA showed increased seed germination and growth compared to control plants (Balki and Padole, 1982). Numerous researches indicated that application of hormones such as auxins may increase the germination ability of seeds and seedling vigour in different plants (Swaminathaan and Srinivasan, 1996; Balestri and Bertini, 2003). Thus it seems that soaking wheat seeds in different auxins before germination counteracted adverse effects of NaCl on adult plant growth.
Significant differences were found in radicle lengths depending on cultivar and salinity and auxin levels. Increasing salt concentrations severely affected radicle elongation (Table 3). The -1.2 and 0.06 MPa NaC1 treatments reduced radicle length in all cultivars in comparison with control. Bewley and Black (1994) suggested that the inhibition of the radicle under salt stress is due to a reduction in the turgor of the radicle cells. The length of radicle was not decreased in high salinity conditions (1.2 MPa) with presence of 1 and 2 mg LG1 auxin for Pishtaz cultivar (Table 3) in comparison with 0 mg LG1 auxin. It seems that Pishtaz cultivar response to auxin that can decrease high salinity effects on some growth traits such as radicle length. These results are consistent with Reed et al. (1998), where, application of auxins accelerates root emergence and enhances root growth in germinated seeds.
There were significant differences in hypocotyle lengths depending on cultivars and salinity and auxin levels. Increasing salt concentrations severely affected on lengths of hypocotyle (Table 3). The -0.6 MPa NaC1 treatment reduced hypocotyle length in Pishtaz cultivar from 12.10 to 9.39 mm while the -1.2 MPa NaC1 reduced hypocotyle length in Mahdavi cultivar from 9.46 to 6.18 mm. Although salinity can induce a rapid reduction in root and shoot growth (Neumann, 1997), radicle length decreases proportionally more than hypocotyle length, causing a reduction in the root/shoot ratio. Salinity significantly reduces the total dry matter and the degree of reduction in total dry matter depending on genotypes and salt concentrations (Pessarakli, 1991). The hypocotyle length increased in salt stress conditions with presence of 1 mg LG1 auxin for all cultivars (Table 3). Thus, all cultivars are appropriate cultivars for auxin usage and auxin presence can be decrease salinity effects on their growth traits.
Increasing salt concentrations severely affected on seedling fresh weight. Mahdavi cultivar under 2 mg LG1 auxin without salinity and Pishtaz cultivar at -1.2 MPa salinity without auxin produced the most (1.45 g) and the least (0.73 g) seedling fresh weight respectively (Table 3). Also increasing salt concentration influenced on seedling dry weight. Mahdavi cultivar at 1 mg LG1 auxin without salinity and Pishtaz cultivar at -1.2 MPa salinity with no auxin produced the most (0.079 g) and the least (0.038 g) seedling dry weights respectively. It seems that reduction in seedling fresh and dry weights is due to decreasing water uptake by seedlings in salt stress presence. This result is similar to the previous study, which showed decreasing seedling fresh weight under 20 mmos cmG1 salinity (Ashraf, 2002). There were not significant differences in radicle dry weight depending on cultivars and auxin levels but, Duncan multiple range test showed radicle dry weight reduction under -0.6 and -1.2 MPa salinity levels (Table 2).
Increasing salt concentrations affected hypocotyle dry weight. Mahdavi cultivar under 1 mg LG1 auxin and Pishtaz cultivar at -1.2 MPa salinity level and 2 mg LG1 auxin produced the most (0.05 g) and the least (0.021 g) hypocotyle dry weight respectively (Table 3).
| Table 3: | Means values (interaction effects) of seed germination traits for three wheat cultivars auxin and salinity stress |
 | |
| Means with similar letter(s) in each trait is not significantly different at 5% probability level according to Duncan's Multiple Range Test | |
| Table 4: | Correlation coefficients among seed germination traits for three wheat varieties cultivars auxin and salinity stress |
 | |
| * and ** significant at 5 and 1%, respectively and ns not significant | |
In both wheat cultivars (Mahdavi and Pishtaz), NaCl decreased hypocotyl dry weight, which is consistent with data obtained by previous authors with other plants (Romer-Aranda et al., 2001).
Auxin increased hypocotyle dry weight in Mahdavi cultivar at high salinity. Pishtaz and Shiraz cultivars are also appropriate cultivars for auxin application to decrease salinity effects on growth traits.
The correlations between each pair of studied traits (seedling dry weight, radicle length, hypocotyl length, seedling fresh weight, radicle dry weight, hypocotyl dry weight and germination percentage) were calculated (Table 4) and demonstrated that germination percentage was only correlated with seedling dry weight (r = 0.76**) and was not correlated with the other measured traits (Table 4). The seedling fresh weight was positively and significantly correlated with seedling dry weight (r = 0.65**), hypocotyl length (r = 0.71**), radicle dry weight (r = 0.51*) and hypocotyl dry weight (0.57*).
There were highly significant (p<0.01) positive correlation between seedling dry weight and the other measured traits except radicle length. Radicle length was negatively correlated (r = -0.069**) with hypocotyl length and was not correlated with radicle and hypocotyl dry weights (Table 4). The hypocotyl length was positively correlated with the hypocotyl dry weight (r = 0.69**), but was not correlated with the radicle dry weight (p>0.05). The correlation was not also significant (p>0.05) between radicle and hypocotyl dry weights (r = 0.40ns).
CONCLUSIONS
Rapid seed germination and stand establishment are critical factors to crop production under salt-stress conditions particularly critical in semi-arid areas where favorable conditions in the seed zone may be brief. In many crop species, seed germination and early seedling growth are the most sensitive stages to salinity stress.
Priming seed with optimal concentrations of plant growth hormones is shown to effectively improve germination as well as growth and yield performance of various crop species under both normal and stress conditions. Growth hormones normally used for seed priming include auxins (IAA, TBA, NAA), gibberellins (GA), kinetin, abscisic acid. In this study demonstrated the positive effects of IAA hormone on some germination traits such as hypocotyl length, seedling fresh and dry weights and hypocotyl dry weight. It seems that moderate levels of auxin (1 mg LG1) could be reduced negative salinity effects on seed germination.
REFERENCES
- Al-Hakimi, A.M.A. and A.M. Hamada, 2001. Counteraction of salinity stress on wheat plants by grain soaking in ascorbic acid, thiamin or sodium salicylate. Biol. Planta, 44: 253-261.
CrossRefDirect Link - Almansouri, M., J.M. Kinet and S. Lutts, 2001. Effect of salt and osmotic stresses on germination in durum wheat (Triticum durum Desf.). Plant Soil, 231: 243-254.
CrossRefDirect Link - Ashraf, M. and M.R. Foolad, 2005. Pre-sowing seed treatment-A shotgun approach to improve germination, plant growth and crop yield under saline and non-saline conditions. Adv. Agron., 88: 223-271.
CrossRefDirect Link - Haidar, M.A., W. Bibi and M.M. Sidahmed, 2003. Response of branched broomrape (Orobanche ramosa) growth and development to various soil amendments in potato. Crop Prot., 22: 291-294.
CrossRef - Balki, A.S. and V.R. Padole, 1982. Effect of pre-soaking seed treatments with plant hormones on wheat under conditions, of soil salinity. J. Indian Soc. Soil Sci., 30: 361-365.
Direct Link - Braccini, A.L., H.A. Ruiz, M.C.L and M.S. Rcis, 1996. Germina��o e vigor de sementes de soja sob estresse h�drico induzido por solu��es de cloreto de s�dio, manitol e polietilenoglicol. Revista Brasileira de Sementes, 18: 10-16.
Direct Link - Chiwocha, S.D.S., A.J. Cutler, S.R. Abrams, S.J. Ambrose, J. Yang, A.R.S. Ross and A.R. Kermode, 2005. The etr1-2 mutation in Arabidopsis thaliana affects the abscisic acid, auxin, cytokinin and gibberellin metabolic pathways during maintenance of seed dormancy, moist-chilling and germination. Plant J., 42: 35-48.
PubMedDirect Link - Ghoulam, C. and K. Fares, 2001. Effect of salinity on seed germination and early seedling growth of sugar beet (Beta vulgaris L.). Seed Sci. Technol., 29: 357-364.
Direct Link - Gulzar, S., M.A. Khan and L.A. Ungar, 2003. Salt tolerance of a coastal salt marsh grass. Commun. Soil Sci. Plant Anal., 34: 2595-2605.
CrossRefDirect Link - Huang, J. and R.E. Redmann, 1995. Salt tolerance of Hordeum and Brassica species during germination and early seedling growth. Can. J. Plant Sci., 75: 815-819.
CrossRefDirect Link - Khan, M.A., I.A. Ungar and A.M. Showalter, 2000. Effects of sodium chloride treatments on growth and ion accumulation of the halophyte Haloxylon recurvum. Commun. Soil Sci. Plant Anal., 31: 2763-2774.
CrossRefDirect Link - Neumann, P., 1997. Salinity resistance and plant growth revisited. Plant Cell Environ., 20: 1193-1198.
CrossRefDirect Link - Reed, D.C., S.J. Holbrook, E. Solomon and M. Anghera, 1998. Studies on germination and root development in the surfgrass Phyllospadix torreyi: Implications for habitat restoration. Aquat. Bot., 62: 71-80.
CrossRefDirect Link - Rekoslavskayal, N.I., O.V. Yurjeva, R.K. Salyaev, S. Mapelli and T.V. Kopytina, 1999. D-tryptophan as IAA source during wheat germination. Plant Physiol., 25: 39-49.
Direct Link - Romero-Aranda, R., T. Soria and S. Cuartero, 2001. Tomato plant-water uptake and plant-water relationships under saline growth conditions. Plant Sci., 160: 265-272.
CrossRefDirect Link - Xue, Z.Y., D.Y. Zhi, G.P. Xue, H. Zhang, Y.X. Zhao and G.M. Xia, 2004. Enhanced salt tolerance of transgenic wheat (Tritivum aestivum L.) expressing a vacuolar Na+/H+ antiporter gene with improved grain yields in saline soils in the field and a reduced level of leaf Na+. Plant Sci., 167: 849-859.
Direct Link