
Tony Hadibarata
Department of Applied Bioscience, Faculty of Agriculture, Ehime University 3-5-7 Tarumi, Matsuyama, Ehime, 790-8566 Japan
Sanro Tachibana
Department of Applied Bioscience, Faculty of Agriculture, Ehime University 3-5-7 Tarumi, Matsuyama, Ehime, 790-8566 Japan
Kazutaka Itoh
Department of Applied Bioscience, Faculty of Agriculture, Ehime University 3-5-7 Tarumi, Matsuyama, Ehime, 790-8566 Japan
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 15 | Page No.: 2535-2543







ABSTRACT
Microbial degradation of Phenanthrene with several fungi screened from nature was conducted to select fungi for the bioremediation of Phenanthrene. Thrichoderma sp. S019, a fungus collected from soil, had the highest rate of degradation on the agar medium containing Phenanthrene. Maximal degradation (72%) was obtained when Trichoderma sp. S019 was incubated for 30 days after the addition of 0.1 mM of Phenanthrene to the liquid medium. Furthermore, the degradation of Phenanthrene was affected by the addition of a carbon source, the addition of a nitrogen source and agitation. Also, 1,2-Dioxygenase and 2,3-Dioxygenase were produced by Trichoderma sp. S019 in a liquid medium. These enzymes play an important role in the metabolism of substrates, revealing a high stereoselectivity for initial dioxygenase and enzymatic hydration since the K-region of phenanthrene was the major site of metabolism. Phenanthrene was indeed degraded by Trichoderma sp. S019 because 1-Hydroxy-2-naphthoic acid, Salicyaldehyde, Salicylic acid and Catechol, considered to be the intermediates in the bioremediation of Phenanthrene, were detected among the reaction products.
PDF Abstract XML References Citation
How to cite this article
Tony Hadibarata, Sanro Tachibana and Kazutaka Itoh, 2007. Biodegradation of Phenanthrene by Fungi Screened from Nature. Pakistan Journal of Biological Sciences, 10: 2535-2543.
DOI: 10.3923/pjbs.2007.2535.2543
URL: https://scialert.net/abstract/?doi=pjbs.2007.2535.2543
DOI: 10.3923/pjbs.2007.2535.2543
URL: https://scialert.net/abstract/?doi=pjbs.2007.2535.2543
INTRODUCTION
Petroleum hydrocarbons are important pollutants found in soil, sediment and marine environments. These compounds are often introduced into the environment through contamination by crude oils, refinery products and harbor and offshore activities, including tanker wreckages. Some of them are toxic, genotoxic, mutagenic and a carcinogenic (Hart, 1996; Heitkam and Cerniglia, 1987). These petroleum hydrocarbon compounds are often introduced into the environment through contamination by crude oils, refinery products and harbor and offshore activities, including tanker wreckages (Morgan and Watkinson, 1989). Consequently, the environment has become highly polluted with different petroleum hydrocarbon compounds. Phenanthrene, a three-ring angular Polycyclic Aromatic Hydrocarbon (PAH), is associated with several carcinogenic compounds (Fawell and Hunt, 1988; Sutherland et al., 1995). Phenanthrene are also considered prototypic PAHs and serve as signature compounds to detect PAH contamination, since their chemical structures are found in carcinogenic PAHs, such as benzo (a) pyrene and benz (a) anthracene. They have also been used as model PAHs to determine factors that affect the bioavailability, biodegradation potential and rate of microbial degradation of PAHs in the environment (Bouchez et al., 1995; Cerniglia, 1992; Kanaly and Harayama, 2000; Sutherland et al., 1995). The metabolism of more complex PAH with four or more rings has been less extensively studied when they are used as a sole carbon source. However, the very low solubility of more complex PAH, in fact, strongly reduces their bioavailability and makes microbial growth and biodegradation difficult. The degradation is also limited by substrate bioavailability in laboratory conditions, where the PAH are supplied in solid form (Boldrin et al., 1993).
The ability of microorganisms to degrade hydrocarbons and facilitate their mineralization by forming more labile organic compounds through the breakdown of intramolecular bonds has been extensively studied. Microbial degradation is a rate-limiting factor in many biogeochemical cycles. As a result, microorganisms have contributed to the development of different bioremediation technologies (Alexander, 1999; Atlas, 1985, Madsen, 1991).
While traditional chemical/physical remediation techniques are currently acceptable from an environmental and economical point of view, there is increasing interest in bioremediation as an alternative approach (Canet et al., 2001). Microorganisms of different origins can be used, nevertheless particular attention has been devoted to indigenous varieties and improving their in situ degradative capability through the optimization of parameters such as temperature, pH, water content and oxygen concentration (Galli, 1999; Hart, 1996). In the last decade, several species of bacteria and filamentous fungi capable of mineralizing or partially degrading PAH with the formation of less harmful products have been identified. Some of the characteristics of filamentous fungi make them better potential degraders than bacteria. In particular, considering the soil habitat, filamentous fungi ramify quickly and are able to grow in environments with low concentrations of nutrients, a low pH and little water (Boonchan et al., 2000; Lamar and White, 2001).
The present study was conducted: (a) to screen, isolate and characterize potential fungi from nature for the degradation of Phenanthrene, (b) to know characteristic of metabolites in the degradation of Phenanthrene, (c) to extract, isolate and characterize the enzymes which play an important role in the degradation of Phenanthrene and (d) to check the optimum treatment for degradation of Phenanthrene.
MATERIALS AND METHODS
Chemicals: Phenanthrene, 1-Hydroxy-2-naphthoic acid and Catechol were purchased from Tokyo Chemical Industry Co. Ltd. (Tokyo, Japan). Malt extract and polypeptone were purchased from Difco. Thin Layer Chromatography (TLC) aluminium sheets (Silica gel 60 F254, 20x20 cm) were obtained from Merk (Darmstadt, Germany). Salicylaldehyde, Salicylic acid, the silica gel used for column chromatography (Wakogel S-1) and all other chemicals were purchased from Wako Pure Chemical Industry Co. Ltd. (Osaka, Japan).
Sample collection and screening of liquid medium: Trichoderma sp. S019 was collected from soil at the Campus of the Faculty of Agriculture, Ehime University and isolated in the Laboratory of Chemistry and Biotechnology for Utilization of Forest Resources, Ehime University, Japan. These experiments were conducted from October 2003 until February 2006. This isolate was selected based on its ability to degrade Phenanthrene in a solid medium containing 20 mL of malt extract with the addition of 1% Phenanthrene and 300 mg L-1 Chloramphenicol or Benomill followed by incubation at room temperature for two weeks and observed daily. Colonies grown on the malt extract agar plates were transferred to a fresh malt extract agar medium to obtain pure strains. The fungus isolated was stored on a malt extract agar plate at 4°C in a refrigerator. Screening of several liquid media was conducted in order to find the optimum medium for degradation of Phenanthrene. The media used were as follows: Mineral Salt Broth (MSB), Harrold’s medium, Potato Dextrose Broth (PDB) and Yeast Malt Extract medium (Arora and Gill, 2001; Makdesi and Beuchat, 1996; Park et al., 2004). Experiments were performed in 100 mL Erlenmeyer flasks containing 20 mL of liquid medium plus Phenanthrene Dissolved in Dimethylformamyde (DMF) to 0.1 mM. In addition, as the strains have different growth rates, the period of incubation varied from 5 to 7 days in order to obtain a similar radial growth and to minimize variation in the starting inoculums. Mycelia plugs of a selected fungus were cut from the outer edge of an actively growing culture on an inoculum plate. Three 5 mm disks obtained by punching out with a cork-borer from the outer edge of an actively growing culture of a particular fungus were inoculated into the flask containing 20 mL of liquid medium supplemented with 0.1 mM of substrates. The flasks were incubated at 25°C. Growth and substrate consumption were determined as 7 day intervals.
Media and culture conditions: Each 5 mm disk of selected fungi were aseptically inoculated into a 100-mL Erlenmeyer flask containing 20 mL of Mineral Salt Broth (MSB) containing (in g L-1 distilled water) : glucose (10), KH2PO4 (2), MgSO4.7H2O (0.5), CaCl2.2H2O (0.1), ammonium tartrate (0.2) and trace element (10 mL) as described by Arora and Gill (2001). The flasks were incubated for 15 and 30 days at 25°C in the dark. After autoclaving for 20 min at 120°C, 1 mM of Phenanthrene dissolved in 1 mL of dimethylformamyde and tween 80 was added to each flask. Control experiments were performed by incubating MSB medium with a test aromatic hydrocarbon without inoculums. All assays were conducted in triplicate. The carbon concentration dependence of Phenanthrene degradation was investigated on media containing seven different carbon sources (glucose, cellobiose, fructose, starch, sucrose, corn steep liquor and xylose) in the concentration range 2-10% of liquid medium. The nitrogen concentration dependence of Phenanthrene degradation was investigated on media containing four nitrogen sources (polypeptone, ammonium tartrate, ammonium nitrate and yeast extract) in the concentration range 2-10% of liquid medium. Agitation at 80 and 120 rpm was conducted to enhance the degradation of Phenanthrene. All media were sterilized by autoclaving at 120°C for 20 min.
Analytical methods: After incubation, the culture broth was blended with ethyl acetate and acidified with 1 N HCl. The filtrate (liquid medium) and residue (fungal body) were separated by filtration and the liquid medium and fungal body was extracted with ethyl acetate, respectively. Each extract was combined and purified by column chromatography using dichlorometane:methanol (19:1) 200 mL. With this method, all substrates initially present in the liquid medium were recovered. The extracts were concentrated and analyzed by gas chromatography-mass spectrometry (GC-MS Shimadzu QP-5050). The amount of substrate was determined using 4-Chlorobiphenyl as an internal standard. GCMS was used with the following conditions: column 30 m in length and 0.25 mm in diameter, helium pressure 100 kPa. The temperature program was started at 80°C, held for 2 min, raised from 80 to 200°C at 20°C min-1, then to 260°C at 7.5°C min-1, then held for 4 min. The flow rate 1.5 mL min-1, interface temperature 260°C and injection volume 1 μL. Degree of degradation was determined by comparison of remaining of phenanthrene between control and samples.
Enzyme assays: 1,2-Dioxygenase and 2,3-Dioxygenase were measured by a modified method of Nakazawa et al. (1970). The malonic acid buffer was added to the culture and centrifuged at 12,000 rpm for 5 min and rapidly washed with 0.1 M NaCl. The cell paste was separated into two parts, one for assay of 1,2-Dioxygenase activity, while another for detection of 2,3-Dioxygenase activity. After putting and stirring for 5 min, the cell paste was carried out and added to DEAE cellulose. The DEAE cellulose was put into the centrifugal separation container and shaken for 5 min. After that, it was put in the refrigerator for 30 min and 50 mL of cell paste was carried out and put into the dialysis film. The reaction was performed at 20°C and after 1 min the numerical value was measured. The enzyme activities in supernatant were determined spectrophometrically at 20°C. All activities were expressed in U, defined as the amount of enzyme required to oxidize 1 μmol of substrate in 1 min.
Detection of metabolites: The Mineral Salt Broth (MSB) medium was prepared as described above. After inoculation of the medium with Trichoderma sp. S019, the culture was pre-incubated by standing for 7 days at 25°C in the dark. Phenanthrene (3.56 mg) dissolved in 100 μL of N, N-Dimethylformamide (DMF) and 10 μL of Tween 80 (1% solution) were added to each culture medium as described above. The incubation was conducted from 7-30 days at 25°C in the dark. The extracts were purified using silica gel column chromatography by successive elution with n-hexane, n-hexane and dichloromethane (1:1 v/v) and methanol. After the vacuum drying of each eluate (100 μL) in a vial, N, O-bis-trimethylsilyl acetamide (40 μL), pyridine (40 μL) and trimethylcholorilane (20 μL) were added, respectively. Trimethylsilylation of the eluate was conducted for 10 min at 80°C without contact with moisture. The Trimethylsilyl (TMS) derivatives of the extract were analyzed by Gas Chromatography (GC) using Shimadzu GC-17 equipped with a TC-1 capillary column (30 mx0.25 mm) ID 0.25 μm using a gradient of 60°C for 2 min, raised to 150°C at 15°C min-1, then raised to 300°C at 25°C min-1 and maintained at 300°C for 6 min. Injector and interface temperatures were 260°C. In order to confirm the intermediates/metabolites of Phenanthrene degradation and to determine the degradation pathway, a Gas chromatograph-mass spectrophotometer, Shimadzu QP5050, was used in this experiment. The conditions for GC-MS analysis consisted of a detector at 1.3 eV, scan intervals of 1 sec and mass range of 50-500.
RESULTS
Isolation and identification of fungi: A total of 124 samples of soil were inoculated on a malt extract agar medium containing Phenanthrene. Among the 124 samples isolated, 87 were selected for further screening by the screening method described above. Seven fungi that grew well on the agar medium were selected among 87 fungi (Table 1). Other isolates were discarded based on their comparatively poor growth on agar medium containing Phenanthrene. As shown in Table 1, one fungus named S109 was found to be most capable of degrading Phenanthrene. Base on macroscopic morphological characteristics, S109 was identified as belonging to Trichoderma sp.
Screening of several liquid media: Mineral Salt Broth (MSB) was chosen as the optimal liquid medium for degradation of Phenanthrene with selected fungi, because MSB showed the highest levels of Phenanthrene degradation with mycelial inoculums equal to 76%, respectively. The degree of degradation of Phenanthrene with Trichoderma sp. S019 in all liquid media was almost the same.
Investigation of degradation of phenanthrene in a liquid medium by selected fungi: The rate of reduction was above 40% for 30 days of incubation with Trichoderma sp. S019. Trichoderma sp. S019 degraded 72 and 31% of Phenanthrene at 0.1 and 1 mM in 30 days, respectively.
| Table 1: | Growth of fungi screened from nature on agar medium containing phenanthrene |
 | |
| Table 2: | Degradation of Phenanthrene by Trichoderma sp. S019 in the liquid medium |
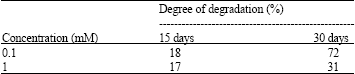 | |
| Table 3: | Effect of nutrients on degradation of Phenanthrene by Trichoderma sp. S019 in the liquid medium |
 | |
There was a simultaneous decrease in the concentration of all compounds during the course of the incubation. Trichoderma sp. S019 gave the highest total reduction at a concentration of 0.1 mM (Table 2).
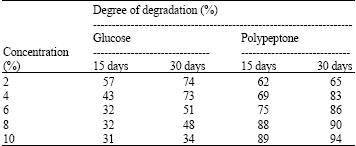
Investigation of effects of carbon and nitrogen sources on degradation of phenanthrene in a liquid medium: Seven carbon sources and four nitrogen sources were added to the Trichoderma sp. S019 cultures. Glucose, sucrose, fructose, cellobiose, starch, corn steep liquor and xylose were used as carbon sources while ammonium tartrate, ammonium nitrate, polypeptone and yeast extract were used as nitrogen sources. After 30 days, glucose and polypepton showed the highest rates of degradation of Phenanthrene, respectively. Addition of glucose and polypeptone in the culture improved the rate to 73 and 64%.
Phenanthrene though easily degraded in the fungal cultures in this study was included in the biodegradation. Trichoderma sp. S019 degraded 74, 73, 51, 48 and 34% of Phenanthrene at 2, 4, 6, 8 and 10% glucose in 30 days, respectively. The maximum Phenanthrene degradation was 73% when Trichoderma sp. S019 was incubated in the culture with 2% glucose in 30 days. Addition of polypeptone was performed in order to know the effect of nitrogen for degradation of the compounds. Trichoderma sp. S019 degraded 65, 83, 86, 90 and 94% of Phenanthrene at 2, 4, 6, 8 and 10% polypeptone in 30 days. The highest rate of degradation was 94% when Trichoderma sp. S019 was incubated in the culture with 10% polypeptone for 30 days (Table 3).
The combination of glucose as a carbon source and polypeptone as a nitrogen source had been used in the dark without shaking. Trichoderma sp. S019 degraded 30, 43 and 36% of Phenanthrene with a combination of glucose and polypeptone of (3:1), (1:1) and (1:3) for 30 days.
| Table 4: | Effect of nutrient mixture ratio on degradation of Phenanthrene by Trichoderma sp. S019 in the liquid medium |
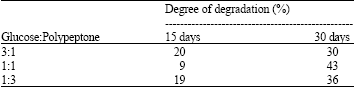 | |
| Table 5: | Effect of agitation on degradation of Phenanthrene by Trichoderma sp. S019 in the liquid medium |
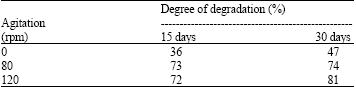 | |
| Table 6: | Effect of nutrients ratio and agitation on degradation of Phenanthrene by Trichoderma sp. S019 in the liquid medium |
 | |
The maximal degradation was 43% when Trichoderma sp. S019 was incubated in the culture with glucose:polypeptone (1:1) for 30 days, respectively (Table 4).
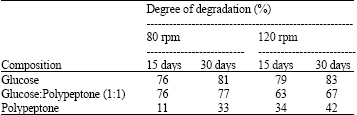
Effects of agitation on the degradation of Phenanthrene in shake-flask cultures were examined in the dark. Trichoderma sp. S019 degraded 74 and 81% of Phenanthrene at 80 and 120 rpm. The maximal degradation rate was 81% when Trichoderma sp. S019 was incubated in the culture with shaking at 120 rpm for 30 days, respectively (Table 5).
Effects of nutrient ratio and agitation on degradation of Phenathrene in the liquid medium by Trichoderma sp. S019 were examined as described above. Trichoderma sp. S019 degraded 83, 67 and 42% of Phenanthrene on addition of glucose, glucose-polypeptone and polypeptone on a 120 rpm rotary shaker for 30 days, respectively. Trichoderma sp. S019 degraded 81, 77 and 32% of Phenanthrene on addition of glucose, glucose-polypeptone and polypeptone on an 80 rpm rotary shaker for 30 days, respectively. The maximal degradation rate of Phenanthrene was 83% when Trichoderma sp. S019 was incubated in the culture with glucose on a 120 rpm rotary shaker for 30 days incubation (Table 6).
Investigation of enzyme activity from selected fungi in the liquid medium: When the fungus was grown in culture for 15 days, the activity of 1,2-Dioxigenase was incubation, the 1,2-Dioxygenase reached 25.3 and 8.3 U for extracted-fungal body and extracted-liquid medium.
| Table 7: | Enzyme activity from Trichoderma sp. S019 in the liquid medium |
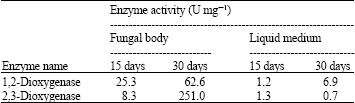 | |
The enzyme of 1,2-Dioxygenase extracted from fungal body shows more activity than that of liquid medium while 2,3-Dioxygenase produced 1.2 and 1.3 U of extracted-liquid medium and extracted-fungal body. After 30 days of incubation, the liquid medium presented more activity than the fungal body. The 1,2-Dioxygenase reached 251 and 62.6 U for extracted-liquid medium and extracted-fungal body. 2,3-Dioxygenase was produced at 6.9 and 0.7 U from extracted-fungal body and extracted-liquid medium, respectively. The maximal enzyme activity was 1,2-Dioxygenase from extracted-liquid medium when Trichoderma sp. S019 was incubated in the culture for 30 days, respectively (Table 7).
Identification of metabolites: To confirm the degradation of Phenanthrene in the liquid medium with fungi, the identification of intermediates was conducted. According to the degradation pathway for Phenanthrene (Pinyakong et al., 2000), 1-Hydroxy-2-naphthoic acid, 2-Hydroxybenzaldehyde, Salicylic acid and Catechol were considered to be major intermediates (Fig. 1). Identification of these compounds in the extracts using the method described in the experimental section was conducted. In the Total Ion Chromatography (TIC) profile, the retention time of these compounds coincided with that of these authentic compounds. The Mass Spectrum (MS) of the trimethylsilyl (TMS) ether of these compounds also coincided with that of the authentic TMS ether of the compound.
The TLC and GC analyses were initially performed to indicate the presence of different intermediates in the degradation pathway combining short-time and long-term extracts of the Phenanthrene-grown culture. The GC-MS studies were then carried out to conclusively prove the presence of these intermediates using short-term extract separately. In this case, the TLC and GC results taken together indicated the presence of 1-Hydroxy-2-naphthoic acid and 2-Hydroxybenzaldehyde. The GC-MS studies were then performed.
Trichoderma sp. S019, grown in MSB with Phenanthrene for 7 days, was able to mineralize Phenanthrene to 1-Hydroxy-2-naphthoic acid. An analysis of the ethyl acetate-extractable metabolites was conducted using GC-MS, under normal conditions. A major peak at 9.6 min, which represented all of the metabolites, was identified as 1-Hydroxy-2-naphthoic acid. The GC retention time, GC-MS properties of the TMS-derivative M+ at m/z 332, fragment ions at m/z 317 (M+-15), corresponding to the respective sequential losses of methyl (-CH3), were identical to those of authentic 1-Hydroxy-2-naphthoic acid (Fig. 3). Several additional Phenanthrene metabolites were also detected by GC-MS analysis of the ethyl acetate-extractable soluble metabolites in short-term incubation. The retention time peak at 7.9 min refers to authentic 2-Hydroxybenzaldehyde. MS analysis of the 2-Hydroxybenzaldehyde produced from 1-Hydro-2-naphthoic acid gave an apparent molecular ion (M+) at m/z 194 for TMS-derivatives and apparent losses of (M+-15) at m/z 179 corresponding to the respective sequential losses of methyl (-CH3) and (M+-16) at m/z 178 corresponding to the respective sequential losses of Oxygen (O) as well as the expected fragment ions at m/z 179 and m/z 73. Based on data obtained above, the compound should be 2-Hydroxybenzaldehyde (Fig. 2).
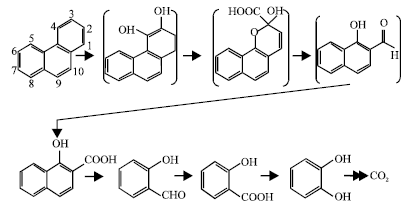 | |
| Fig. 1: | A proposed pathway for the degradation of Phenanthrene by Trichoderma sp. S019 with reference to the results of Pinyakong et al. (2000) |
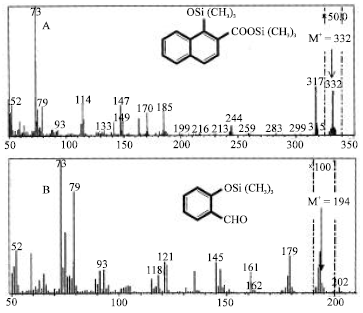 | |
| Fig. 2: | Mass spectra of the trimethylsilyl (TMS) ether of the short-term incubation product from bioremediation of Phenanthrene with Trichoderma sp. S019 screened from nature. (A) 1-Hydroxy-2-napththoic acid, produced from bioremediation of Phenanthrene, (B) 2-Hydroxybenzaldehyde, produced from bioremediation of Phenanthrene |
 | |
| Fig. 3: | Mass spectra of the trimethylsilyl (TMS) ether of the long-term incubation product from bioremediation of Phenanthrene with Trichoderma sp. S019 screened from nature. (A) Salicylic acid, produced from bioremediation of Phenanthrene, (B) Catechol, produced from bioremediation of Phenanthrene |
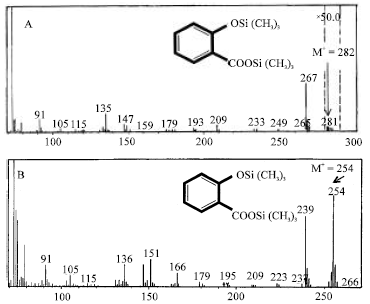
After incubation of Trichoderma sp. S019 in MSB medium for 15 days, the ethyl acetate extract was verified to contain one metabolite by analytical TLC. The extract was partially purified by Wakogel S-1 silica gel column chromatography followed by TLC. The TLC results revealed that the metabolite was present in the fraction containing dichloromethane:methanol (1:1). The fraction was pooled and analyzed by GC-MS. We detected one peak which had a GC retention time of 6.9 min. On GC-MS analysis, this metabolite had a molecular ion (M+) at m/z 282 and fragment ion (M+-15) at 267, representing a probable loss of CH3. This mass spectral fragmentation pattern suggested that this metabolite was salicylic acid (Fig. 4). Long-term incubation was conducted with Trichoderma sp. S019 for 30 days. By the GC analysis of metabolites after silylation, we detected one TMS-derivative peak. The mass spectrum of the major compound having a retention time of 5.7 min exhibited a molecular ion (M+) at m/z 254 and fragment ion at (M+-15) at 239 representing a probable loss of CH3..
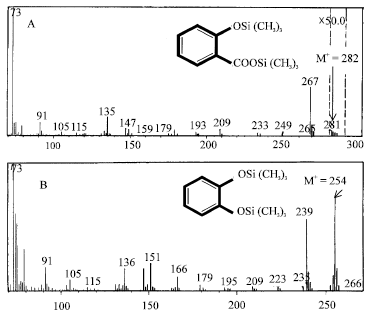 | |
| Fig. 4: | Mass spectra of the trimethylsilyl (TMS) ether of the long-term incubation product from bioremediation of Phenanthrene with Trichoderma sp. S019 screened from nature. (A) mass spectrum of Salicylic acid, produced from bioremediation of Phenanthrene, (B) mass spectrum of Catechol, produced from bioremediation of Phenanthrene. |
This metabolite was identical as Catechol based on a comparison with the authentic compound (Fig. 3).
DISCUSSION
In this study, we isolated a filamentous fungus, Trichoderma sp. S019 that is able to degrade Phenanthrene. Chavez-Gomez et al. (2003) reported that Phenanthrene could be removed from soil by co-cultures of bacteria and fungi including Trichoderma viride. Trichoderma sp. S019 utilized PAH as a sole carbon source for growth. There have been relatively few studies on the growth of Trichoderma sp. S019 with Phenanthrene as a carbon source. In the present experiment, when Phenanthrene was provided an usually long period, 30 days, of incubation was required to reach very low levels of Phenanthrene. This could be due to the fact that Phenanthrene was used at very high concentrations coupled with it being the only exogenous carbon source provided. Chavez-Gomez et al. (2003) have reported a strain of Trichoderma viride, which grows in a medium containing Phenanthrene. The fungus could degrade phenanthrene in sugarcane bagasse pith by around 30% after 18 days of incubation. The degradation rate of Phenanthrene by Trichoderma viride in sugarcane bagasse pith was lower that of Trichoderma sp. S019 in the liquid medium. Another previous study has shown that Trichoderma harzianum and Trichoderma pseudokoningii isolated from soil have the ability to degrade pyrene in a liquid medium (Ravelet et al., 2000).
Polypeptone as a nitrogen source, when added to the MSB medium, remarkably increased the rate of degradation of Phenanthrene. The Phenanthrene concentration reached a low level within 30 days of incubation. An interesting observation recorded in the present study was the apparent link between the decrease in the Phenanthrene concentration and the level of glucose in the medium. This result is similar to that of Ezzy and Lynch (2005) who found that degradation of substrate decreased after glucose was added to the medium. The explanation given by them, for this phenomenon is that the loss is not due to the action of microbes, wherein glucose reacts with free Phenanthrene. However, in the control set up without any inoculation of the microbial culture, the concentration of glucose remained almost constant throughout the study with a negligible loss of glucose recorded. This indicates that glucose was indeed used by the fungi for a long period growth with Phenanthrene. The ability of microorganisms to degrade PAH is a function of the number of aromatic rings in the compound. An increase in the concentration of substrate decreased the degradation rate by Trichoderma sp. S019. It was suggested that the growth of the fungi was inhibited at a higher concentration of substrate. The rate of Phenanthrene’s degradation in the absence and presence of glucose and polypepton was totally different. There was a significant increase in biomass.
In agreement with reports of the deleterious effect of agitation on enzyme production, agitation of growing shake-flask cultures on a rotary shaker at 120 rpm resulted in the formation of mycelial pellets. This result is similar to that of Collina et al. (2005) that the oxygen concentration is directly dependent on the air flow rate. On the one hand, stirring increases the contact between the reagents (substrate, oxygen and biomass), thus enhancing mass transfer and, as a consequence, the biodegradation rate. On the other hand, agitation increases the contact surface between the slurry phase and the gas phase in the bioreactor, thus enhancing the loss by volatilization of oxygen and substrates and decreasing the biodegradation rate.
The degradation of Phenanthrene was a complicated process because many enzymes and different catabolic pathways are involved and many intermediates were produced, accumulated and consumed when the degradation proceeds. The initial 1,2-Dioxygenase and 2,3-Dioxygenase was responsible for the first step in the aerobic degradation of polyaromatic compounds, catalyzing the hydroxylation of the substrate to the corresponding cis-dihydrodiol (Pinyakong et al., 2000). Meyer et al. (1999) explained that the catalytic meta-cleavage of catechol by 1,2-Dioxygenase and 2,3-Dioxygenase seems to be the most common pathway in the subsequent steps of PAH degradation. We traced the activities of these two important enzymes in this research.
Many PAHs contain a bay region and a K-region. The bay- and K-regions, which can be formed metabolically, are highly reactive both chemically and biologically (Cerniglia and Gibson, 1980b). Biodegradation, both prokaryotic and eukaryotic, requires the presence of molecular oxygen to initiate enzymatic attack on the PAH rings (Cerniglia, 1984). Trichoderma sp. S019 was cultivated in the presence of individual aromatic hydrocarbons. Phenanthrene, a low molecular weight PAH with three benzene rings, is easily metabolized by a wide variety of fungi, including Trichoderma sp. S019 and representatives of other fungal groups. The data on identified metabolites, together with the finding that Trichoderma sp. S019 can utilize catechol, have been used to propose a degradation pathway for Phenanthrene. According to the pathway, Phenanthrene was degraded by Trichoderma sp. S019 via dioxygenation at 3, 4-position followed by meta-cleavage. As ring-cleavage products are unstable, cis-3,4-Dihydroxy-3,4-dihydrophenanthrene and 3,4-Dihydroxyphenanthrene can be oxidized to 1-Hydroxy-2-naphthoic acid, 2-Hydroxybenzaldehyde, Salicylic acid and Catechol, strongly suggesting that the initial dioxygenase used for Phenanthrene degradation by Trichoderma sp. S019 has a broad substrate specificity making studies on the function of initial dioxygenase in this strain interesting. 1-Hydoxy-2-naphthoic acid and Salicylic acid are the conventional intermediates, as reported earlier by Samanta et al. (1999). No early intermediates of Phenanthrene degradation could be detected. Filamentous fungi hydroxylate PAHs as a prelude to detoxification, whereas most bacteria oxidize PAHs as a prelude to ring fission and carbon assimilation (Cerniglia and Gibson, 1980a; Resnick et al., 1996). Similar to bacteria, fungal metabolism of PAHs is also highly region and stereoselective (Cerniglia, 1984; Cerniglia and Gibson, 1980a, b; Cerniglia et al., 1992).
Microbial degradation of PAHs is thought to be the major process involved in effective site bioremediation. The number of PAH compounds now known to be biodegraded has increased; however, the number of isolates of microorganisms that can completely mineralize high molecular weight PAHs, such as chrysene, is limited. Four and five-ring PAHs are very recalcitrant due to their low solubility and the resonance energies of their structures.
ACKNOWLEDGMENTS
We thank Mr. Djarwanto, Forest Product Technology Research and Development Center, Ministry of Forestry, Bogor, Indonesia for identification of the fungus Trichoderma sp. S019.
REFERENCES
- Arora, D.S. and P.K. Gill, 2001. Comparison of two assay procedures for lignin peroxidase. Enzyme Microb. Tech., 28: 602-605.
CrossRef - Atlas, R.M., 1995. Petroleum biodegradation and oil spill bioremediation. Mar. Pollut. Bull., 31: 178-182.
CrossRefDirect Link - Boonchan, S., M.L. Britz and G.A. Stanley, 2000. Degradation of high-molecular-weight polycyclic aromatic hydrocarbons by defined fungal-bacterial cocultures. Applied Environ. Microbiol., 3: 1007-1019.
Direct Link - Bouchez, M., D. Blanchet and J.P. Vandecastelle, 1995. Degradation of polycyclic aromatic hydrocarbons by pure strains and by defined strain associations: Inhibition phenomena and cometabolism. Applied Microbiol. Biotechnol., 43: 156-164.
CrossRefDirect Link - Canet, R., J.G. Birnstingl, D.G. Malcolm, J.M. Lopez-Real and A.J. Beck, 2001. Biodegradation of PAHs by native micro flora and combinations of white rot fungi in a coal-tar contaminated soil. Bioresour. Technol., 76: 113-117.
Direct Link - Cerniglia, C.E., 1992. Biodegradation of polycyclic aromatic hydrocarbons. Biodegradation, 3: 351-368.
CrossRefDirect Link - Chavez-Gomez, B., R. Quintero, F. Esparza-Garcia, A.M. Mesta-Howard and F.J. Zavala Diaz de la Serna et al., 2003. Removal of phenanthrene from soil by co-cultures of bacteria and fungi pregrown on sugarcane bagasse pith. Bioresour. Technol., 80: 177-183.
Direct Link - Collina, E., G. Bestetti, P. Di Gennaro, A. Franzetti, F. Gugliersi, M. Lasagni and D. Pitea, 2005. Naphthalene biodegradation kinetics in an aerobic slurry-phase bioreactor. Environ. Int., 31: 167-171.
CrossRef - Hart, S., 1996. In situ bioremediation: Defining the limits. Environ. Sci. Technol., 30: A398-A401.
Direct Link - Kanaly, R.A and S. Harayama, 2000. Biodegradation of high-molecular-weight polycyclic aromatic hydrocarbons by bacteria. J. Bacteriol., 182: 2059-2067.
CrossRefDirect Link - Makdesi, A.K. and L.R. Beuchat, 1996. Improved selective medium for enumeration of benzoate-resistant, heat-stressed Zygosaccharomyces bailii. Food Microbiol., 13: 281-290.
CrossRefDirect Link - Meyer, S., R. Moser, A. Neef, U. Stahl and P. Kampfer, 1999. Differential detection of key enzyme of polyaromatic hydrocarbon degrading bacteria using PCR and gene probes. Microbiology, 145: 1731-1741.
PubMedDirect Link - Pinyakong, O., H. Habe, N. Supaka, P. Pinpanichkarn and K. Juntongjin et al., 2000. Identification of novel metabolites in the degradation of phenanthrene by Sphingomonas sp. Strain P2. FEMS Microbiol. Lett., 191: 115-121.
PubMedDirect Link - Ravelet, C., S. Krivobot, L. Sage and R. Steiman, 2000. Biodegradation of pyrene by sediment fungi. Chemosphere, 40: 557-563.
CrossRefDirect Link - Resnick, S.M., K. Lee and D.T. Gibson, 1996. Diverse reaction catalyzed by naphthalene dioxygenase from Pseudomonas sp. strain NCIB 9816. J. Ind. Microbiol. Biotechnol., 17: 438-457.
Direct Link - Samanta, S.K., A.K. Cakraborti and R.K. Jain, 1999. Degradation of phenanthrene by different bacteria: Evidence for novel transformation sequence involving the formation of 1-naphthol. Applied Microbiol. Biotechnol., 53: 98-107.
PubMedDirect Link