
Hossein Hassanpour
Department of Basic Sciences, Department of Clinical Sciences, College of Veterinary Medicine,
Sharekord University, Sharekord, Iran
Pejman Mirshokrai
Department of Clinical Sciences, College of Veterinary Medicine,
Sharekord University, Sharekord, Iran
Abolfazl Shirazi
Department of Clinical Sciences, College of Veterinary Medicine,
Sharekord University, Sharekord, Iran
Atefe Aminian
Department of Basic Sciences,
Department of Clinical Sciences, College of Veterinary Medicine,
Sharekord University, Sharekord, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 14 | Page No.: 2374-2378







ABSTRACT
The aim of this study is to investigate the effects of NO on sperm motility of ram. After incubation of normozoospermic samples for 160 minutes in the presence of L-arginine (NOS substrate ), SNP (NO donor), L-NAME (non-selective nitric oxide inhibitor) and packed erythrocytes (NO scavenger), sperm motility assessed in four grades (A, B, C, D). In this study, L-arginine and SNP non-significantly increased progressive motility at low concentration and significantly decreased progressive motility at high concentration. L-NAME and packed erythrocytes dose- dependently decreased progressive sperm motility. It is concluded that sperm motility of ram physiologically depend on nitric oxide action although excess generation of nitric oxide by sperm or exogenous NO could not increase sperm motility and it seems that excess NO provides toxic condition to decrease sperm motility.
PDF Abstract XML References Citation
How to cite this article
Hossein Hassanpour, Pejman Mirshokrai, Abolfazl Shirazi and Atefe Aminian, 2007. Effect of Nitric Oxide on Ram Sperm Motility In vitro. Pakistan Journal of Biological Sciences, 10: 2374-2378.
DOI: 10.3923/pjbs.2007.2374.2378
URL: https://scialert.net/abstract/?doi=pjbs.2007.2374.2378
DOI: 10.3923/pjbs.2007.2374.2378
URL: https://scialert.net/abstract/?doi=pjbs.2007.2374.2378
INTRODUCTION
Nitric oxide (NO) is a gaseous free radical with short-half life that functions as a potent cell signaling, effector and vasodilator molecule in numerous tissues. NO is synthesized from a guanidino nitrogen atom of the essential amino acid L-arginine by a family of isoenzymes known as the nitric oxide synthases (NOS) (Pacher et al. 2007) in the presence of oxygen and the electron donors Nicotinic Acid Adenine Dinucleotide (NAD), Flavin Adenine Dinucleotide (FAD) or flavin mononucleotide (FMN) with L-citrulline occurring as a by-product. Three major NOS isoforms, which share a common basic structural organization and requirement for substrate cofactors for enzymatic activity, have been described. Endothelial (eNOS or NOS III) and neuronal (nNOS or NOSI) isoforms are constitutive, Ca2+ /calmodulin-activated and inducible isoform (iNOS or NOSII) is Ca2+¯ independent and induced by many inflammatory factors such as cytokines and lipopolysaccharide (LPS) ( Michel and Feron, 1997). Following the recognition of NO as a mediator of penile erection (Burnett, 2006), NOS protein and activity have been demonstrated both in male and female reproductive organs (McCann et al., 1999; Telfer et al., 1995), suggesting an involvement of NO in the physiology of reproduction. Evidence has been reported that NO can also be generated by spermatozoa. An immunoreactivity for NOS was observed in mouse, human (Herrero et al., 1996) and bull sperm (Meiser and Schulz, 2003).
It has been shown that human spermatozoa with normal motility, express significant eNOS protein while low motility spermatozoa exhibit aberrant patterns of eNOS expression (O’Brayan et al., 1998). In semen collected from different subjects, a positive correlation was observed between the concentration of NO and percentage of immotile spermatozoa (Rosselli et al., 1995). The present study sought to investigate NO involvement in the motility of ram spermatozoa using activator, scavenger and inhibitor of NO or its enzymes.
MATERIALS AND METHODS
Chemicals: Synthetic oviductal fluid buffered with HEPES (SOF-HEPES) medium, L-arginine, Sodium nitroprusside (SNP), and NG-nitro-L-arginine methyl ester (L-NAME) were purchased from Sigma chemical company (St Louis, Mo, USA). Packed erythrocytes obtained from ram after removing of plasma and washing by saline.
Animals and management: The study was performed on Bakhtiari sheep in a farm of Agricultural Research Center (Shooli, Sharekord, Iran) at October and November. The rams were kept as a single flock in a semi-extensive production system. Bakhtiari rams were with mean weight of 95±5 kg and body condition scores (BCS) of 3.0 to 4.0 (scale 0-5).
Semen processing and Preparation of sperm suspension: Semen was collected using an artificial vagina from twenty Bakhtiari rams. Generally one ejaculate per ram was collected on a daily basis. All ejaculates were evaluated using light microscopy within 30-45 minutes after collection (keeping in 37°C). Ejaculates were only used if the volume was = 0.5 mL and had a sperm concentration = 2x109 spermatozoa/ml and = 60% motile cells. After measuring the sperm concentration (Neubauer chamber, 100 x magnifications) by light microscope, Motile spermatozoa were harvested by the swim-up technique (37°C for 1h in air atmosphere) (Parrish et al., 1986) using SOF-HEPES medium .The presence of round cells (spermatogonia, spermatocytes, spermatides and leucocytes) were minimal (less than 1x106 mL-1) in all sperm samples after swim-up technique in final suspension. After swim-up, the motile sperm-rich fraction was centrifuged at 600xg for 10 min at room temperature and after light microscopy evaluation again, sperm concentration was adjusted to approximately 5 x108 cells mL-1 by adding SOF-HEPES medium. Aliquots of sperm suspension in SOF-HEPES medium, 500 μL each, containing 2 x107 cells, were incubated for 120 min in presence of (0.1, 0.5, 0.7 μM) SNP, ( 1, 10, 100μM) L-NAME, (0.1, 1, 10 mM) L-arginine and (1, 10 μL mL-1) Packed erythrocytes separately and an equal volume of SOF-HEPES medium (controls). Then motility was evaluated by light microscopy according to WHO manual recommendation (WHO, 1999) at 37°C. Spermatozoa were graded: rapid progressive motility, grade A; slow or sluggish progressive motility, grade B; non-progressive motility, grade C; or immotility, grade D. At least 200 spermatozoa in five different microscopic fields were evaluated for each sample.
Statistical analysis: Results are expressed as mean±SEM that is obtained from at least 7 experiments in each group. The difference between experimental groups was tested by Student’s t-test using SPSS-14 package. P-values less than 0.05 were considered statistically significant.
RESULTS
Effect of L-arginine in sperm motility: After incubation of sperm suspension by different concentrations of L-arginine, it was found that concentrations of 0.1 and 1 mM caused sperm motility grades A and C to be increased and grade D decreased, although these variations weren’t significant (p<0.05). Concentration of 10 mM conversely decreased grade A (and increased grade D significantly (p<0.05). Variations of this concentration in grade B and C weren’t significant (Fig. 1).
Effect of SNP in sperm motility: SNP in the concentration of 0.1 μM non-significantly increased sperm motility at grades A and C and decreased at grades B and D.
Concentration of 0.5 μM increased grade B and D, and decreased grade C. SNP at the concentration of 0.7 μM significantly decreased sperm motility at grade A (15.1%) and increased at grade D (16.7%), in comparison with their control groups (p<0.05). Decreasing of sperm motility at grades B and C at concentration of 0.7 μM weren’t significant (Fig. 2).
Effect of L-NAME in sperm motility: L-NAME at concentrations of 1 and 10 μM non-significantly decreased grade A and increased grade D.
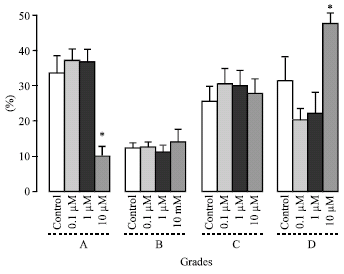 | |
| Fig. 1: | Effect of L-arginine on sperm motility (A: fast progressive motility, B: slow progressive motility, C: non-progressive motility and D: immotility), in different concentrations. Values are means±SE. *p<0.05 from corresponding control |
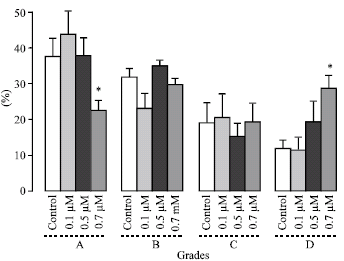 | |
| Fig. 2: | Effect of SNP on sperm motility (A: fast progressive motility, B: slow progressive motility, C: non-progressive motility and D: immotility) in different concentrations. Values are means±SE. *p<0.05 from corresponding control |
Variations in grade B and C were minimum. Concentration of 100 μM caused sperm motility grades A (15.3%) significantly (p<0.05) and B non-significantly to be decreased and grade C non-significantly and D (10.2%) significantly increased in compared with their controls (p<0.05) (Fig. 3).
Effect of packed erythrocytes on sperm motility: Packed erythrocytes at concentration of 1 μL mL-1 significantly (p<0.05) decreased grade A (7.1%) and non-significantly increased grade D.
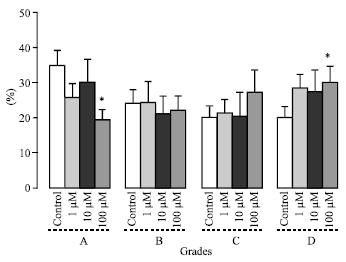 | |
| Fig. 3: | Effect of L-NAME on sperm motility (A: fast progressive motility, B: slow, progressive motility, C: non-progressive motility and D: immotility) in different concentrations. Values are means±SE. *p<0.05 from corresponding control |
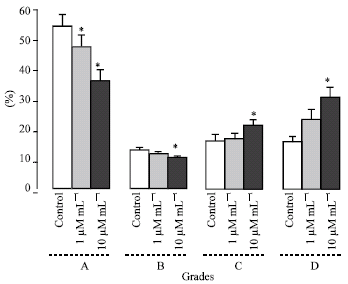 | |
| Fig. 4: | Effect of packed erythrocytes on sperm motility (A: fast progressive motility, B: slow progressive motility, C: non-progressive motility and D: immotility) in different, concentrations. Values are means±SE. *p<0.05 from corresponding control |
Variations in grade B and C were minimum. Concentration of 10μL mL-1 showed grades A (18.2%) and B (2.5%) to be reduced and grades C (5.4%) and D (15%) enhanced. These variations were significant when compared to their control groups (p<0.05) (Fig. 4).
DISCUSSION
NO is a molecule of great biological significance and has long been considered to play an important role in sperm physiology such as sperm chemotaxis, sperm motility, sperm-egg interaction and spermatogenesis (Revelli et al., 2002). Many studies suggest that spermatozoa could represent one of the targets of NO, and a role for NO in controlling the function of male gametes during their maturation and migration in the genital tracts is conceivable. In fact, human and rodent sperm have been reported to be influenced by exogenous NO, as far as motility, viability, and metabolism are concerned (Herrero et al., 1994; Hellstrom et al., 1994; Weinberg et al., 1995). NO could even participate in the mechanisms leading to fertilization, as in bovine sperm it increases capacitation and acrosomal reactivity (Zamir et al., 1995).
In this study, at the first it is found that effects of L- arginine on sperm motility are dose- dependent in vitro, and low concentrations of L- arginine increasingly affect on sperm motility, while high concentration of L-arginine conversely decrease. Srivastava et al. (2006) determined that NO is synthesized from L-arginine by the enzyme Nitric Oxide-Synthase (NOS) present in spermatozoa, and they suggested a possible participation of NO and NOS in arginine action. In vitro studies have shown that low concentrations of NO enhance the motility of mouse (Herrero et al., 1994) and human (Zhang and Zheng, 1996) spermatozoa and medium/high concentrations of NO decreasing sperm motility (Rosselli et al., 1995). It is tempting to speculate in our condition, small amounts of NO are generated by L-arginine that neutralizes free radicals which inhibit sperm motility. In contrast, excessive generation of NO by high concentration of L-arginine can cause sperm toxicity as well as reduce sperm motility by contributing to the formation of peroxynitrite, a highly toxic anion of peroxidation (Levonen et al., 2001). Ratnasooriya and Dharmasiri (2001) also demonstrated that L-arginine caused a dose-dependent impairment of hyperactivated motility in vitro, which can obviously inhibit fertilizing potential of sperm. However, this data confirm our findings.
Present results showed that SNP as a No donor, could affect sperm motility increase at low concentration. In contrast, its high concentration declined sperm motility, these findings as similar as Rosselli et al. (1998), Nobunaga et al. (1996) and Weinberg et al. (1995) results, also strongly suggest that exogenous NO is beneficial at low concentration for ram sperm motility and is harmful at high concentration.
At this study, L-NAME as nitric oxide synthase inhibitor and packed erythrocytes as nitric oxide scavenger dose-dependently decreased progressive motility and increased immotile sperm. These data provide evidence that motility of ram spermatozoa is regulated autocrinally by nitric oxide. A study by Lewis et al. (1996) also suggested the presence of eNOS and nNOS in the human spermatozoon that regulates (increases) sperm motility in an autocrine fashion. and they demonstrated that, as compared to normozoospermic samples, the expression of eNOS and concentrations of NO generated were lower in asthenozoospermic samples, suggesting that decreased endogenous NO may influence sperm motility and hence fertilization.
Taken together, the present study showed that sperm motility of ram physiologically is affected nitric oxide action although excess generation of nitric oxide by sperm or exogenous NO couldn’t increase sperm motility and it seems that excess NO provide toxic condition to decrease sperm motility.
ACKNOWLEDGMENTS
This study had been supported by the funds granted by the Applied Research Centre, Vice Chancellor for Research of Sharekord University. Authors would like to thank Dr. Ahmadi for his assistance during this research.
REFERENCES
- Burnett, A.L., 2006. Nitric oxide in the penis-science and therapeutic implications from erectile dysfunction to priapism. J. Sex. Med., 3: 578-582.
Direct Link - Hellstrom, W.J.G., M. Bell, R. Wang and S.C. Sikka, 1994. Effects of sodium nitroprusside on sperm motility, viability and lipid peroxidation. Fertil. Steril., 61: 1117-1122.
Direct Link - Herrero, M.B., E. Cebral, M. Boquet, J.M. Viggiano, A. Vitullo and M.A. Gimeno, 1994. Effect of nitric oxide on mouse sperm hyperactivation. Acta Physiol. Pharmacol. Ther. Latinoam., 44: 65-69.
Direct Link - Herrero, M.B., S.P. Martinez, J.M. Viggiano, J.M. Polak and M.F. de Gimeno, 1996. Localization by indirect immunofluorescence of nitric oxide synthase in mouse and human spermatozoa. Reprod. Fertil. Dev., 8: 931-934.
Direct Link - Levonen, A.L., R.P. Patel, P. Brookes, Y.M. Go and H. Jo et al., 2001. Mechanisms of cell signaling by nitric oxide and peroxynitrite: From mitochondria to MAP kinases. Antioxid. Redox. Signal., 3: 215-229.
Direct Link - Lewis, S.E.M., E.T. Donnelly, E.S.L. Sterling, M.S. Kennedy, W. Thompson and U. Chakravarthy, 1996. Nitric oxide synthase and nitrite production in human spermatozoa: Evidence that endogenous nitric oxide is beneficial to sperm motility. Mol. Hum. Reprod., 2: 873-878.
Direct Link - McCann, S.M., C. Mastronardi, A. Walczewska, S. Karanth, V. Rettori and W.H. Yu, 1999. The role of nitric oxide in reproduction. Brazil. J. Med. Biol. Res., 32: 1367-1379.
Direct Link - Meiser, H. and R. Schulz, 2003. Detection and localization of two constitutive NOS isoforms in bull spermatozoa. Anat. Histol. Embryol., 32: 321-325.
Direct Link - Michel, T. and O. Feron, 1997. Nitric oxide synthases: Which, where, how and why? J. Clin. Invest., 9: 2146-2152.
Direct Link - Pacher, P., J.S. Beckman and L. Liaudet, 2007. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev., 87: 315-424.
CrossRefPubMedDirect Link - Parrish, J.J., J.L. Susko-Parrish, M.L. Leibfried-Rutledge, E.S. Critser, W.H. Eyestone and N.L. First, 1986. Bovine in vitro fertilization with frozen-thawed semen. Theriogenology, 25: 591-600.
CrossRefDirect Link - Ratnasooriya, W.D. and M.G. Dharmasiri, 2001. L-arginine, the substrate of nitric oxide synthase, inhibits fertility of male rats. Asian J. Androl., 3: 97-103.
Direct Link - Revelli, A., D. Ghigo, F. Moffa, M. Massobrio and I.T. Kaspa, 2002. Guanylate cyclase activity and sperm function. Endocrine Rev., 23: 484-494.
Direct Link - Rosselli, M., R.K. Dubey, B. Imthurn, E. Macas and P.J. Keller, 1995. Effects of nitric oxide on human spermatozoa: Evidence that nitric oxide decreases sperm motility and induces sperm toxicity. Hum. Reprod., 10: 1786-1790.
Direct Link - Rosselli, M., R.J. Keller and R.K. Dubey, 1998. Role of nitric oxide in the biology, physiology and pathophysiology of reproduction. Hum. Reprod. Update., 4: 3-24.
Direct Link - Srivastava, S., P. Desai, E. Coutinho and G. Govil, 2006. Mechanism of action of L-arginine on the vitality of spermatozoa is primarily through increased biosynthesis of nitric oxide. Biol. Reprod., 74: 954-958.
Direct Link - Telfer, J.F., F. Lyall, J.E. Norman and I.T. Cameron, 1995. Identification of nitric oxide synthase in human uterus. Hum. Reprod., 10: 19-23.
Direct Link - Weinberg, J.B., E. Doty, J. Bonaventura and A.F. Haney, 1995. Nitric oxide inhibition of human sperm motility. Fertil. Steril., 64: 408-413.
Direct Link - Zamir, N., D. Barkan, N. Keynan, Z. Naor and H. Breitbart, 1995. Atrial natriuretic peptide induces acrosomal exocytosis in bovine spermatozoa. Am. J. Physiol., 269: E216-E221.
Direct Link - Zhang, H. and R.L. Zheng, 1996. Possible role of nitric oxide on fertile and asthenozoospermic infertile human sperm functions. Free Radic. Res., 25: 347-354.
Direct Link