
Md. Moniruzzaman Sarker
Department of Chemistry, Biology and Marine Science, Faculty of Science, University of the Ryukyus, 1 Senbaru, Nishihara City, Okinawa 903-0213, Japan
Badrun Nesa
Department of Chemistry, Biology and Marine Science, Faculty of Science, University of the Ryukyus, 1 Senbaru, Nishihara City, Okinawa 903-0213, Japan
Md. Sarwar Jahan
Department of Zoology, University of Rajshahi, Bangladesh
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 1 | Page No.: 23-31







ABSTRACT
The egg type of Lymnaea acuminata was determined as iso-lecithal and the cleavage is spirally holoblastic type. The development of L. acuminata was observed in details. Uncleaved zygote just after laying was found to contain a relatively yolk-free zone, the animal pole and the yolk-rich region, the vegetal pole. No polar bodies were present in eggs examined immediately after they had been laid. The first polar lobe and polar body were extruded out from the zygote within 15-25 min. These were reabsorbed after 12-15 min. The formation of the second polar lobe was followed within the next 48 min. The first cleavage division occurred about 115-130 min after the formation and re-absorption of the second polar lobe, retaining for 7-10 min. The 2-celled embryos underwent the second equal division of cleavage the within next hour and the embryos reached the 4-cell stage. At the end of the 5th h, the 4-celled embryos underwent the third cleavage and the cleavage was horizontal (i.e., equatorial). At the end of the 9th h, the embryos at the 8-celled stage reached the 16-celled stage by the 4th cleavage. The 4th spirally cleaved embryos usually underwent fifth and sixth cleavages within 24-26 and 27-29 h of incubation, respectively. After 2-3 days of incubation, the developing embryos attained the trochophore stage. At the beginning of the 4th day of incubation, embryos became slightly elongated, curved foot muscle and shell gland were developed through the extension of velum and the embryos turned into the early veliger. At the beginning of the seventh day, the miniature snail possessed all the structures found in a newly hatched individual. Interaction between water physico-chemical parameters and some breeding parameters have been observed.
PDF Abstract XML References Citation
How to cite this article
Md. Moniruzzaman Sarker, Badrun Nesa and Md. Sarwar Jahan, 2007. Embryonic Developmental Ecology of Freshwater Snail Lymnaea acuminata (Lymnaeidae: Gastropoda). Pakistan Journal of Biological Sciences, 10: 23-31.
DOI: 10.3923/pjbs.2007.23.31
URL: https://scialert.net/abstract/?doi=pjbs.2007.23.31
DOI: 10.3923/pjbs.2007.23.31
URL: https://scialert.net/abstract/?doi=pjbs.2007.23.31
INTRODUCTION
Lymnaeids are distributed worldwide (Godan, 1983). This snail is of immense importance from the viewpoint of economy. They are the source of animal protein and act as the intermediate hosts of trematode parasites, the causative agent of helminth diseases. Moreover, they play an important part in the aquatic ecosystems. Like all other animals in the ecosystem, the distribution and abundance of freshwater snails are also influenced by the environmental components, such as, water qualities, aquatic vegetation, soil substratum, depth of water, temperature, turbidity, pH Dissolved Oxygen (DO), hardness, etc. (Mukherjee, 1986; Raut, 1986; Jahan and Rahman, 1996). In most living things a new generation begins with the fertilized eggs or zygote, a cell formed by the union of the ovum from the female parent and the sperm from the male. The word zygote, derived from Greek, means yoked or jointed together, reminding us that each of the parents has a role in determining the characteristics of the new individual, its size, shape, physiologic attributes and behavioural patterns. The totality of the process whereby these characteristics are achieved and their changes throughout adulthood and senescence, eventually leading to death, are called development (Saunders, 1970). Brahmachary (1989) stated that embryonic development is a ceaseless process till eclosion, which involves several aspects such as zygote metabolism, cleavage, blastulation, gastrulation and organogenesis. The morphological and behavioural aspects of the development of Lymnaea have been well documented (Morril, 1982; Voronezskaya et al., 1999). DeWitt (1954) studied the reproduction, embryonic development and growth in the pond snail Physa gyrina Say. He mentioned the mechanism of copulation, oviposition, breeding parameters like eggs, egg capsule, cleavage and larval development and growth of P. gyrina. Cumming (1993) worked on the reproduction and variable larval development of an ectoparasitic snail, Turbonilla sp. He described the chronology of larval development and mentioned only one larval stage, veliger in the snail species. Okusu (2002) made an observation on the embryogenesis and development of Epimenia babai.
Barraud (1957), Bayne (1968), Suliman et al. (1987), Wijsman et al. (1987), Lara et al. (1988), Pasic and Ambrasic (1990), Presing (1993), Ferguson et al. (1993), Herman et al. (1994) and Rondelaud and Dreyfuss (1996) described the reproduction and embryology of the different species of Lymnaea. The freshwater snails such as Lymnaea develop through what correspond to trochophore and veliger stage with in transparent egg capsules (Mescheryakov, 1990; Morril, 1982; Raven, 1966). The capsules, while conveniently permitting observations of growing embryos, act as a diffusion barrier and prevent passage of large molecules. But still now, detailed work on embryology of L. acuminata with factor affecting the development is completely lacking. In Bangladesh only Sarker and Jahan (1998), Sarker et al. (2000) and Sarker (2002) worked on the embryonic development of L. stagnalis with some details information.
Embryological development is a very sensitive period for any species. It is highly influenced by the changes of ecological factors. For good snail farming one should know the tolerance level and mode of adaptability to severe changes of ecological factors during the developmental period for the test species. For the scientific management of ecologically important species like L. acuminata, clear and adequate knowledge on its development and the ecological factors responsible for the process deserve proper pursuit. Keeping this idea in mind the present investigation was undertaken to study the sequential events of the embryonic development of L. acuminata with the role played thereon by physico-chemical components like total hardness, magnesium hardness, calcium hardness, pH and dissolve Oxygen of water.
MATERIALS AND METHODS
The experiment was conducted at the Ecological research laboratory of the Department of Zoology, University of Rajshahi during the period from May 2002 to December 2003.
In the laboratory a colony of L. acuminata was maintained in filtered, dechlorinated and copper-free tap water (artificial pond water; APW) at 29.57±0.17°C on a 12:12 light: dark cycle. The snails were fed a diet consisting of lettuce, carrot and sweet potato. Aquatic vegetations like Nymphaea, Scirpus, Lemna and Pistia were supplied also for resting and egg laying. To study the embryogenesis and embryokinesis as well as organogenesis of L. acuminata freshly laid egg capsules were collected from the stock culture with the help of a camel hairbrush regularly. The length and width of eggs and developing embryos in the eggs were measured using occulometer in compound microscope (Eagle Instrument Co. Model-180). A batch of 14 freshly land egg capsules were kept separately five glass beakers (500 mL) containing tap water. The experiment was replicated ten times. The developing embryos were studied in vivo with the help of a compound microscope throughout the incubation period and changes were recorded regularly.
Preparation of permanent slide: For preparing permanent slides, eggs incubated for various durations were processed applying the following consecutive procedures: Kept in 50% ethyl alcohol for 30 min, Kept in 70% ethyl alcohol for 40 min, Stained with 1% aqueous solution of Bismark-Brown followed by alcoholic eosin and Mounted with cover slip in D.P.X. Photographs were taken by useing microphotographic apparatus (Model-Leica-III).
Growth: Average geometric growth rates were calculated for two series of developing embryos using the formulation of Simpson and Roe (1939):
Where:
| Yo | = | Initial magnitude |
| Yt | = | Magnitude at time t |
| t | = | Time measured at intervals of 1 day |
Physicochemical properties of water: A total of five sets of experiment were designed for the determination of incubation period and hatchability in respect of physical and chemical parameters of water used for rearing. Physicochemical parameters viz., temperature, pH, dissolved oxygen, total hardness were measured through Hach Kit (Model: FF 1A).
RESULTS
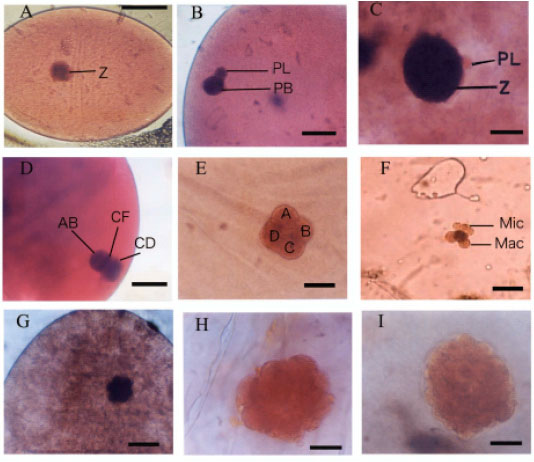
Egg type: The eggs of L. acuminata are isolecithal. The ovum was located eccentrically within the egg (Fig. 1A) and remained in this position until the embryos reached the trochophore stage. Uncleaved zygotes just after laying were found to contain a relatively yolk-free zone, the animal pole and the other yolk-rich region named as vegetal pole.
 | |
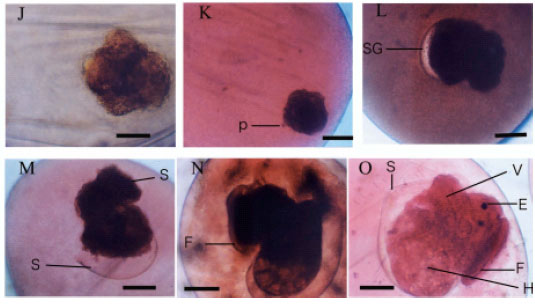 | |
| Fig. 1: | Micrograph of the early embryological development of Lymnaea acuminata until hatching . A. fertilized egg of just after spawning. B. First polar body. C. Second polar body. D. First cleavage; two-cells (AB/CD) stage. E. Four cells stage of developing embryo (A-D). F. Eight cells stage of developing embryo. Mic; Micromere, Mac; Macromere G. Sixteen cells stage of developing embryo. H. 32- cells of developing embryo. I. 64-cells stage of developing embryo. J. Gastrula stage of developing embryo. K. Trochophore stage. L. Early veliger stage. M Veliger stage. N. Late veliger stage. O. Before hatching. Z = Zygote, CF = Cleavage furrow, E = Eye, F = Foot, H = Heart, P = Protroch, PB = Polar body, PL = Polar lobe, SG = Shell gland, S = Shell, V = Velum. Scale bars = 100 μm |
The series of embryological events, during the experimental period took place at the prevailing room temperature (29-31°C).
Cleavage: No polar bodies were present in eggs examined immediately after they had been laid. The first polar lobe and polar body were extruded out from the zygote within 15-25 min (Fig. 1B). These were reabsorbed after 12-15 min. The formation of the second polar lobe (Fig. 1C) was followed within the next 48 min. The first cleavage division occurred about 115-130 min after the formation and re-absorption of second polar lobe, retaining for 7-10 min. Cellular division was rapid in the animal pole than that in the vegetal pole. Due to the presence of little yolk, spiral holoblastic cleavage occurred in the zygote of L. acuminata. The cleavage of L. acuminata was characterized by very early occurrence of a cleavage cavity. Already at the 2 cell stage a wide cavity was formed between the two blastomeres and this was repeated in the succeeding cleavages. The cleavage furrow at first cut very deeply into the zygote (Fig. 1D), so that the daughter cells finally touched each other at one point only.
The first division was typical, it passed through the animal-vegetal pole, although it might be inclined somewhat (Fig.1D) due to absorption of first and second polar lobes and first polar body which ultimately gave rise to a bit larger blastomere (CD) with the smaller one (AB). The two-celled embryos underwent the second equal division of cleavage within the next hour and the embryos reached the 4-cell stage producing four large blastomeres called macromeres, designated arbitrarily as A, B, C and D (Fig. 1E).
At the end of 5th h, the 4-celled embryos underwent the third cleavage and the third cleavage was horizontal (i.e., equatorial). The third cleavage was also frequently unequal, so that the cells of the upper tier (the micromere) were smaller than those of the lower tier (the macromeres) (Fig. 1F). At this stage a second polar body appeared and was reabsorbed frequently. At the end of the 9th h, the embryos of the 8-celled stage reached the 16-celled stage (Fig. 1G) by the 4th cleavage. The 4th spirally cleaved embryos usually underwent fifth and sixth cleavages within 24-26 and 27-29 h of incubation respectively (Fig. 1H and 1I).
Blastulation: The exact process of blastulation could not be recognized. We saw no blastocoel in the developing embryos.
Gastrulation: In the present experiment, in L. acuminata, gastrulation started at an early phase of development after 30-32 h of incubation, when the embryo contained more than 64 cells (Fig. 1J). The macromeres and their descendants, situated at the centre of the vegetal half, were changed in shape. The external surface of the macromeres became reduced whereas the inner part got widened so that they became wedge shaped.
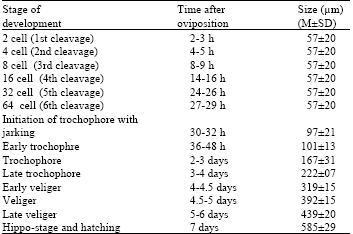
Trochophore stage: During the late hours (33-35) of gastrulation a very slow jerking movement of the developing embryos were noticed to attain the early trochophore stage (Table 1). Through the following hours the embryos took a bit elongated shape and within 36-48 h of incubation the embryos developed with mouth, velum or prototroch, head vesicle, etc. (Fig. 1K). At that time the embryos gained the revolving movement and attained the complete trochophore stage. The next 24 h of incubation period was passed for the attainment of adequate development as the late trochophore. Timing, stage and size of developing embryos with physico-chemical parameters of water used in the experiment are presented in Table 1.
Veliger stage: At the beginning of the 4th day of incubation embryos became slightly elongated, had curved foot muscle and shell gland developed through the extension of velum and the embryos turned into the early veliger (Fig. 1L). After five days bilobed foot anlagen of the veliger larva and the head foot region sharply became demarcated from the visceral mass (Fig. 1M). A pair of tentacles bearing darkly pigmented eyes at their bases was developed after six days of incubation. The heartbeat was clearly observed during the 6th day and the embryo exhibited considerable coordination of movement (Fig. 1N and O). Periodically (every 15-20 sec) the visceral mass was contracted toward the head-foot region.
| Table 1: | Duration, developmental stages and sizes of developing embryos of L. acuminata |
 | |
| Table 2: | Measurement of growth of two groups of embryos with the average geometric growth rate of L. acuminata |
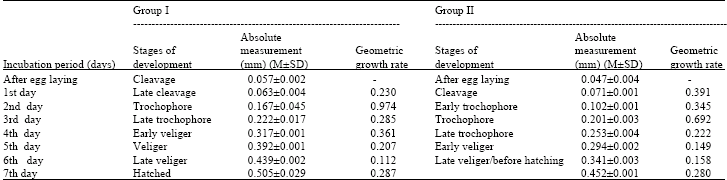 | |
| The eggs in group I were laid 12 h earlier than those in group II (n = 10) | |
Hatching: The young snails escaped from the egg by first rasping through the egg membrane. Perforation of the membrane is aided by the snail periodically thrusting its foot against the wall. The process was completed within 20-30 min. After hatching, the empty egg membrane collapsed. Outside the egg, the snail moved freely in the jelly of the egg capsule and escaped out through a hole made by itself or through an opening made by another snail previously.
Juveniles: At the beginning of the 7th day, the miniature snail possessed all the structures found in a newly hatched individual. The foot and tentacles characteristic of Lymnaeidae were formed and a thin transparent shell covered the mantle. The growing embryonic snails nearly filled the egg by the end of the seven days. Hatching usually began on the seventh and was completed by the beginning of the eighth day.
Embryonic growth of L. acuminata: For the single-celled zygote to produce a multicellular organism, a number of mitotic divisions must occur in rapid succession. After 2 h of oviposition the first cleavage was noticed in L. acuminata zygote, which produced a 2-celled stage. During cleavage the size and shape of the embryo is retained the same, while the cleaved cells, or blastomeres, became smaller at each division and this trend was observed from 2-cell to 64-cell stages. Table 2 contains the stages of development, timing and size of the developmental stages from oviposition through hatching of juvenile L. acuminata.
Embryonic growth of L. acuminata was studied in the peak period of the breeding season in June and July 2003. Embryos were measured at daily intervals from oviposition until hatching and the values are presented in Table 2. It was observed that embryonic growth of L. acuminata follows the same general pattern found in other Metazoa. There is no appreciable increase in total size of the embryo during cleavage; as the total number of cells increase, they become smaller. Measurable increase in the mass of embryonic tissue follows gastrulation and subsequent development of the structures characterizing the trochophore and Table 2.
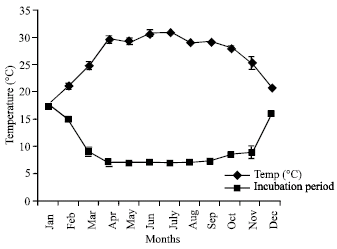 | |
| Fig. 2: | Effect of temperature on the Incubation period of L. acuminata |
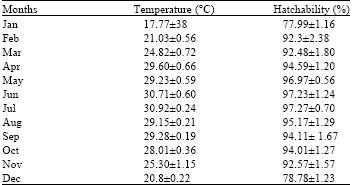
| Table 3: | Effects of temperature on the hatchability of L. acuminata |
 | |
In veliger stage many of the organs found in the adult made their initial appearance. Following this period, the rate of growth decreased.
Incubation period and hatchability in respect of physicochemical properties of water: Effects of various physicochemical characters of water on the incubation period and hatchability have been presented in Fig. 2,
| Table 4: | Prevailing physical and chemical factors of water and two breeding parameters of L. acuminata with correlation matrix among them (Pearsons correlation) |
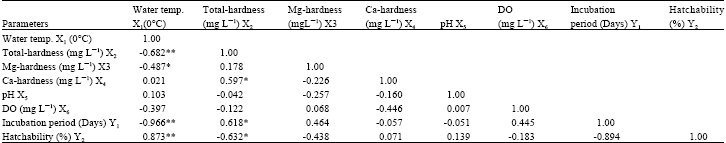 | |
| **Correlation is significant at the 0.01% level, * Correlation is significant at the 0.05% level | |
Table 3 and 4. The highest water temperature was recorded in July (30.92±0.24°C) and the lowest in January (17.77±0.38°C).
The minimum and maximum incubation periods of 6.9±0.12 and 17.5±0.86 days were July and January, respectively. The minimum and maximum hatchabilities of 77.99±1.16 and 97.28±1.24% were recorded in January and June, respectively. Suddenly after February, due to high temperature, the occurrence of dissolved oxygen decreased reversely with the water temperature.
Pearson’s correlations among some water parameters viz., water temperatures (X1), total hardness (X2), magnesium hardness (X3), calcium hardness (X4), dissolved oxygen (X6) and breeding parameters, viz., incubation period (Y1) and hatchability (Y2), have been presented in Table 4. Data indicate that the water temperature was positively correlated with calcium hardness, pH and hatchability (p<0.01) but negatively correlated with total hardness (p<0.05), magnesium hardness (p<0.01), dissolved oxygen and incubation period (p<0.01). Total hardness was positively correlated with magnesium hardness, calcium hardness (p<0.05) and incubation period (p<0.05) but negatively correlated with pH, dissolved oxygen and hatchability. Magnesium hardness is positively correlated with dissolved oxygen, incubation period but negatively correlated with calcium hardness, pH and hatchability. Calcium hardness was positively correlated with hatchability but negatively correlated with pH, dissolved oxygen and incubation period. pH was positively correlated with dissolved oxygen and hatchability but negatively correlated with incubation period. Lastly, dissolved oxygen is positively correlated with incubation period but negatively correlated with hatchability. Incubation period was negatively correlated with hatchability.
DISCUSSION
Gilbert (1991) stated that the eggs of most molluscs are isolecithal except cephalopods. Cleavage is a progressive subdivision of zygote by mitotic cell division. The pattern of cleavage was holoblastic, spiral which has been supported by Verdonk and Biggelaar (1983). The available data strongly suggest that in molluscan eggs at least three supercompartments exist, viz the animal pole, the vegetal pole and the cortical region. In eggs of Lymnaea a distinct animal pole plasm became visible about 1 h before the first cleavage (Dohmen, 1983). Certain spirally cleaving embryos (mostly in Molluscs and Annelids) extrude a bulb of cytoplasm immediately before first cleavage. This protrusion is called the polar lobe (Gilbert, 1991).
Verdonk and Biggelaar (1983) mentioned that opisthobrnch and the pulmonate cleavages with polar lobe have no documentation. The finding of Gilbert (1991) has been supported by the present observation made on L. acuminatai with some differences noticed regarding the time of the formation of polar lobe and polar body. Wilson (1904) concluded that the first lobe extruded out as a polar lobe immediately before the first cleavage in Dentalium. He experimentally showed that in the trifoil stage removing the polar lobe, produced a trochophore larva, which produced incomplete larva completely lacking mesodermal organs e.g., muscles, mouth, shell gland and foot. Clement (1952) also stated that after removal of the polar lobe at the first cleavage, a radially symmetrical embryo was formed, which failed to develop some adult structures. In the present investigations it was noticed that before the first cleavage both polar lobe and polar body were formed at the same time and after their re-absorption, the second polar lobe was formed and re-absorbed again. After this the first cleavage division occurred. A second polar body developed which disappeared between the 3rd and 4th cleavages. The same phenomena of appearance and disappearance of the polar body after the 3rd cleavage were reported by and Gilbert (1991) in the snail Ilyanassa.
The pattern of cleavage in Bithynia, Dentilium and Ilyanassa were described by Browder (1980). He stated that spiral cleavage is found in some flatworms, the nemerteans and the molluscs (except cephalopods). The spiral pattern is apparent at the eight-cell stage. The arrangement of blastomeres in a spirally cleaving embryo is caused not by rearrangement of cells after cleavage but rather by the oblique orientation of spindles prior to cleavage. The first division is typical: it passes through the animal-vegetal pole axis, although it may be somewhat inclined. The spindles at the second division are perpendicular to the first. However, unlike in radial cleavage in which they would also be perpendicular to the main axis, these spindles are inclined and in opposite directions to one another. The third cleavage is equatorial (i.e., horizontal) as radial cleavage, but the spindles are oblique rather than parallel to the major axis. Consequently, the cleavage planes are shifted, causing the cells of the animal quartet to lie in the spaces between the lower cells. The third cleavage is also frequently unequal, so that the cells of the upper tier (the micromeres) are smaller than those of the lower tier (the macromeres). At successive cleavages the micromeres divide equally, producing additional quartets of micromeres, which are arranged in tiers stacked below pre-existing ones. Gilbert (1991) informed about cleavage and Wilson (1904) described the processes of spirally cleavage in annelids, turbellarian flatworms, nemerteans and molluscs. Browder (1980) described the cleavage pattern of molluscs in details. Unfortunately, nobody mentioned timing of the cleavage period in L. acuminata. Raven (1948) described only the timing of the first two cleavages of L. stagnalis at 20°C.
The present study revealed that cleavage would have been completed within 24-26 h of incubation at 29-31°C. The first cleavage division occurred about 115-130 min after the formation of the second polar body. The first division was typical. It passed through the animal-vegetal pole. The two-celled embryos underwent the second equal division of cleavage within the next hour and the embryos reached the 4-celled stage. At the end of 5th h, the 4-celled embryos underwent the third cleavage and the third cleavage was horizontal (i.e., equatorial) which was frequently unequal and produced an upper tier, the micromeres and the lower tier, macromeres. But (Desrosiers et al., 1996) stated that in giant scallop Placopecten magellanicus, first division was unequal and produced larger CD blastomere than the AB though the cleavage time was similar with present investigation. At the end of the 9th hour of incubation, the embryos of the 8-celled stage reached the 16-celled stage by the 4th cleavage. The 4th spirally cleaving embryos usually underwent fifth and sixthcleavages and later on (128-cell stage) reached the gastrula stage through a true blastula stage. This supports the statement of Verdonk and Bigglaar (1983), that gastrulation in Lymnaea started when the embryo consisted of 120 cells and with the advancement of gastrulation the blastopore became narrow and assumed the form of a slit. The blastulae so produced by the cleavage had no blastocoel and were steroblastulae. These have also been supported by Gilber (1991).
L. acuminata laid egg capsules containing eggs with zygotes 57±20 μm long. Within the next 2-28 h, the length of the cleaving zygote remained constant. However, there was an increase in the number of smaller blastomeres in the developing embryos. Such a phenomenon was observed by Cumming (1993) in the opisthobranch, Turbonilla. During the late hours of gastrulation at 28-32 h post oviposition, when the developing embryo reached a length of 97±21, a very slow jerking movement of the embryo was noticed. The elongated embryo with developing mouth, velum and head vesicle gained the revolving movement at the end of 2-3 day’s of incubation and reached the trochophore stage. The trochophore was 167±31 μm in length. The highest geometric growth rate was observed at the trochophore stage which indicates that the most rapid growth take place at the trochophore stage in the pulmonate snails. This finding is similar to those obtained by DeWitt (1954) and Sarker and Jahan (1998). Development in basal groups of gastropod molluscs includes a dramatic movement that has intrigued embryologists and evolutionary biologists for over a century. The process is called ontogenetic torsion (Page, 2002). But in the present investigation the process of torsion observation was lacking. Further more work is necessary to explore the ontogenetic torsion of the test species.
Significant difference between development in L. acuminata and Helix were observed mainly in the general body structure. Thus the velum was present in L. acuminata and rudimentary type in Helix (Victor et al., 2001). Ranjah (1942) stated that breeding season and the incubation period varies with the nature of the environment. When the temperature varied between 90-100°F, the incubation period was 10-40 days and due to a constant and heavy rainfall when the temperature came down to 70 to 80°F, the incubation period was about three weeks. Raut (1981) concluded that the rate of breeding and fluctuation in the number of snails, viz., Viviparus bengalensis, L. acuminata, I. exustus and Acrostoma variabilis were governed by temperature, rainfall and food. He also added that favourable water temperature with rainfall were essential for breeding in snails so that the population increased at an appreciable rate. In the present investigations, the maximum incubation period was recorded as 17.50±0.86 days in January when the minimum temperature was 17.77±0.38°C. But the minimum incubation period of 6.90±0.12 days was recorded at 30.92±0.24°C in July. The minimum and maximum hatchabilities were 77.99±1.16 and 97.27±0.75% in January and July, respectively. These results are similar to those stated above.
Present study confirms that, this mollusc is easy to handle and culture and is available in various quantities and stages of embryonic development throughout the year are similar with previous studies (Canton and Slooff, 1977). In conclusion, knowledge of optimal conditions for successful embryonic development will useful for biochemical and cytological studies of early development in L. acuminata, for achieving maximal productivity in snail hatcheries, as well as for genetic manipulations, such as production of triploids.
Different important stages of development and their timing are the essential finding for future researchers.
Temperature influences the total hardness, magnesium hardness and dissolved oxygen negatively but calcium hardness and pH positively, which indicate that the chemical properties of water are determined by the physical environmental component, temperature. From the above experimental results it can be concluded that temperature played vital role controlling the development of L. acuminata. Aziz and Raut (1996) stated that hatching success of eggs varied with pH and salinity grades it is obvious that factor have significant influence to alter the beneficial effects of temperature, to certain extent though they did not consider the hardness of water. Raut (1989) stated that the development of L. luteola was highly influenced by pH of water and beyond the range of 6.0-9.5, eggs of snail were not hatched. Present results with L. acuminata development agree with those reported for oyster Crssostrea virginica, the clam Mercenaria mercenaria (Calabrese and Davis, 1966) and giant scallop Placopecten magellanicus (Desrosiers et al., 1996), which shows successful embryonic development at pH values of 6.75-8.75, 7.00-8.75 and 8.0-8.5, respectively.
Temperature, calcium hardness and pH negatively influenced the incubation period that means they reduced the incubation period and enhanced hatchability. But total hardness, magnesium hardness and dissolved oxygen increased the incubation period and restricted the hatchability through incomplete or slow development of the embryos. The presence of calcium hardness in water is also an important factor for the proper development of L. acimunata. In this experiment we observed the interaction of some ecological factors and breeding parameters but not different range of this factors. Further work is needed to explore the optimum range of this factors for proper development and growth of cultured snail.. The data obtained from this study suggest that L. acimunata has wide range of ecological factors which indicate their wide range of survibality. Further experimentation, however is clearly needed to refine methods for snail controlling development in aquaculture and to understand the mechanism by which factors strongly operate in the L. acuminata. The observations made, open a new avenue for future workers to establish developmental mechanism and snail culture farms. However, much more research is solicited in this line.
ACKNOWLEDGMENTS
The authors are grateful to the Chairman, Department of Zoology, University of Rajshahi, for providing laboratory facilities and to Professor Dr. Ataur Rahman Khan, Department of Zoology, Rajshahi University and to Professor Dr. Makoto Tsuchiya, Department of Chemistry, Biology and Marine Science, University of the Ryukyus, Japan, for his painstaking, meticulous job of going through the manuscript and constructive criticism.
REFERENCES
- Aziz, M.A. and S.K. Raut, 1996. Thermal effect on the life-cycle parameter of the medically important snail species Lymnaea (Radix) luteola (Lamarck). Mem. Inst. Oswald. Cruz., 91: 119-128.
Direct Link - Desrosiers, R.R., J. Desilets and F. Dube, 1996. Early developmental events following fertilization in the giant scallop Placopecten magellanicus. Can. J. Fish. Aquat. Sci., 53: 1382-1392.
Direct Link - Ferguson, G.P., A.W. Pieneman, R.F. Jansen and A.T. Maat, 1993. Neuronal feedback in egg-laying behaviour of the pond snail Lymnaea stagnalis. J. Exp. Biol., 178: 251-259.
Direct Link - Godan, D., 1983. Pest Slugs and Snails, Biology and Control. Springer-Verlag, Berlin, Germany, ISBN: 0387118942, Pages: 445.
Direct Link - Herman, P.M., A. Maat and R.F. Jansen, 1994. The neural control of egg laying behabiour in the pond snail Lymnaea stagnalis: Motor control of shell turning. J. Exp. Biol., 197: 79-99.
Direct Link - Page, L.R., 2002. Ontogenetic torsion in two basal gastropods occurs without shell attachments for larval retractor muscles. Evol. Dev., 4: 212-222.
Direct Link - Presing, M., 1993. Influence of an insecticide K-othrine, on the reproduction and mortality of the pond snail (Lymnaea stagnalis L.). Arch. Envion. Cont. Toxi. USA., 25: 387-393.
Direct Link - Victor, N.I. and M. Balamban, 2001. Ontogenesis of the snail, Helix aspersa: Embryogenesis timetable and ontogenesis of GABA-like immunoreactive neurons in the central nervous system. J. Neurocytol., 30: 73-91.
Direct Link - Pasic, M. and M. Ambrasic, 1990. Bioelectrical activity of Lymnaea stagnalis L. nervous system during embryonal development. Zool. Jahr. Abteil. Allgem. Zool. Physiol. Tier., 94: 181-188.
Direct Link