
Ali Karimi Goudarzi
Department of Basic Sciences, College of Veterinary Medicine,
Islamic Azad University, Karaj Branch, Karaj, Iran
Hossein Hassanpour
Department of Basic Sciences, College of Veterinary Medicine, Sharekord University, Sharekord, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 12 | Page No.: 2091-2094







ABSTRACT
The aim of present study was to characterize nitric oxide production via nitrite measurement as its stable metabolite in the different species of spermatozoa (human, bull and ram). Sperm samples were fractionated on the basis of density with the use of a three-layer Percoll gradient. Spermatozoa collected from the various percoll gradient interfaces (50/70 and 70/95%). Results showed that nitrite is produced by human, bull and ram spermatozoa, but in the same sample, there were no significant difference between high density (with higher percentage of motility) sperm subpopulation and low density (with lower percentage of motility) sperm subpopulation Therefore, it seems that spermatozoa with different motility in the one sperm population have the same production of NO in physiological condition that is different from pathological condition (e.g., in the subfertile men).
PDF Abstract XML References Citation
How to cite this article
Ali Karimi Goudarzi and Hossein Hassanpour, 2007. In vitro Production of Nitrite by Low and High Density Sperm Subpopulations of Human, Bull and Ram. Pakistan Journal of Biological Sciences, 10: 2091-2094.
DOI: 10.3923/pjbs.2007.2091.2094
URL: https://scialert.net/abstract/?doi=pjbs.2007.2091.2094
DOI: 10.3923/pjbs.2007.2091.2094
URL: https://scialert.net/abstract/?doi=pjbs.2007.2091.2094
INTRODUCTION
Nitric Oxide (NO) is a molecule utilized throughout the animal kingdom as a signaling or toxic agent between cells. NO is now known to be produced by various cells in different organs, including smooth muscle cells, mesangial cells, neurons, platelets, hepatocytes, macrophages, fibroblasts and epithelial cells. NO regulates smooth muscle cell tone, platelet aggregation and adhesion, cell growth, apoptosis, neurotransmission and injury as well as infection-induced immune reactions (Pacher et al., 2007; Lincoln et al., 1997). Because these processes are also associated with the biology, physiology and pathophysiology of various reproductive processes, it is highly likely that NO plays an important role in reproduction (McCann et al., 1999).
NO is synthesized universally from L-arginine and molecular oxygen by an enzymatic process that utilizes electrons donated by NADPH. The NO synthase (NOS) enzymes convert L-arginine to NO and L-citrulline via the intermediate N-hydroxy-L-arginine. NO is generated either by the constitutively expressed enzymes NOS-1 and NOS-3 or the induced enzyme NOS-2. NOS-1 and NOS-3 are activated in response to physiological stimuli that trigger an intracellular Ca2+ signal; they produce NO rapidly and transiently at low levels. NOS-2 is not expressed in resting cells but is induced by immunological stimuli such as bacterial lipopolysaccharide (LPS) or cytokines such as IL-1, TNF-α or IFN-γ (Michel and Feron, 1997; Coleman, 2001).
In the past studies, NO has been recognized as a molecule that importantly regulates the biology and physiology of the reproductive system and not surprisingly then, there is now strong evidence supporting a role for NO in modeling sexual and reproductive functions in mammalian species. Following the recognition of NO as a mediator of penile erection (Burnett, 2006), NOS protein and activity have been demonstrated both in male and female reproductive organs (Burnett et al., 1995; Telfer et al., 1995), suggesting an involvement of NO in the physiology of reproduction. Evidence has been reported that NO can also be generated by spermatozoa. An immunoreactivity for NOS was observed in mouse, human (Herrero et al., 1996) and bull sperm (Meiser and Schulz, 2003). The aim of present study is to characterize nitric oxide production (via nitrite measurement as a stable metabolite of nitrite oxide) in the different species of spermatozoa (human, bull and ram) and to examine the relation between NO synthesis and sperm quality.
MATERIALS AND METHODS
Chemicals: Hepes-buffered Tyrode albumin lactate pyruvate (Hepes-TALP) medium components (114 mM NaCl, 3.1 mM KCl, 0.3 mM NaH2PO4, 2.1 mM CaCl2, 0.4 mM MgCl2, 2 mM NaHCO3, 0.2 mM sodium pyruvate, 10 mM sodium lactate, 10 mM Hepes, 5 mg mL–1 BSA and 0.7 mg L–1 Pen/Strep), Sulfanilamide 1% and N-1-napthylethylenediamine dihydrochloride (NED) 0.1% [under acidic (phosphoric acid) conditions] and Heparin were purchased from Merck Chemical Co. (Bubendorf, FRG). Lowry reagents, Earle’s Balanced Salts Solution (EBSS) and Percoll solution were from Sigma Chemical Co. (St. Louis, MO).
Preparation of human semen samples: Semen samples were obtained by masturbation after 3-5 days of sexual abstinence from 10 donors. All samples were allowed to liquefy for at least 30 min at 37°C.
Preparation of cauda epididymal spermatozoa from bull and ram: After preparation of ten cauda epididymides from abattoir and transport to laboratory close to ice, were cut by a sterile scalpel, then 0.5 mL of the released epididymal contents were transferred to 35 mm Petri dishes containing 2 mL of equilibrated Hepes-TALP (for ram sperm) or Hepes-Heparin (10 μg mL–1)-TALP (for bull sperm) medium. All dishes containing samples were incubated in a humidified atmosphere of 5% CO2 in air at 37°C.
Discontinuous percoll gradient: All samples were evaluated for sperm concentration, motility and morphology according to the WHO guidelines (World Health Organization, 1992). Only specimens with normal parameters (World Health Organization, 1992) were used in the experiments. Sperm samples were fractionated on the basis of density with the use of a three-layer Percoll gradient (de Lamirande and Gagnon, 1991). Spermatozoa collected from the various percoll gradient interfaces (50/70 and 70/95%) were washed by centrifugation at 500 x g for 20 min in Hepes-TALP (for ram sperm), Hepes-Heparin (10 μg mL–1)-TALP (for bull sperm) or BSA-EBSS medium (for human sperm).
Measurement of nitrite: Suspensions of human, bull and ram spermatozoa (20x106 cells in 1 mL) obtained from the two percoll gradient interfaces were incubated for 24 h in the their individual mediums. Nitrite production was measured by adding 0.15 mL of supernatant to 0.15 mL of Griess reagent (Green et al., 1982) and after 10 min incubation at 37°C in the dark, absorbance at 540 nm was measured using a light spectrophotometer. A blank was prepared for each experimental condition in the absence of cells and its absorbance was subtracted from that obtained in the presence of cells. Nitrite concentration was expressed as nanomoles of nitrite produced in 24 h mg–1 cell protein. Mass protein of cells was measured by method of Lowry et al. (1951).
Statistical analysis: Statistical analyses were conducted using SPSS version 14.0 for Windows (SPSS Institute Inc.). All results are represented as mean±SEM. Comparisons were made using ANOVA. Statistical significance was regarded when p<0.05.
RESULTS
Low-density (50/70% Percoll interface) and high-density (70/95% Percoll interface pooled with a 95 layer) sperm subpopulations were obtained by centrifugation on discontinuous Percoll gradients. The low and high-density sperm subpopulation of human, bull and ram incubated for 24 h in their culture media produced orderly nitrite in the amounts of 74.9±11.8 and 42.5±7.4 (human), 356.9±56.6 and 280.4±50 (bull), 35.7±5.6 and 46.7±1.9 (ram) nanomole/24 h/cell protein. These amounts between low and high density sperm subpopulations were nonsignificant in the each species while between bull and other species were significant (p<0.05) (Fig. 1). Low density sperm subpopulation of bull produced nitrite 4.75-fold more than Human and ten fold more than ram low density sperm subpopulation. High density sperm subpopulation of bull released nitrite 6.6-fold more than Human and six fold more than ram high density sperm subpopulation.
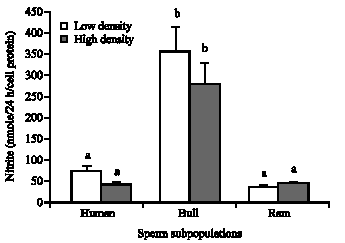 | |
| Fig. 1: | Comparison of the nitrite production between Low Density (LD) and High Density (HD) sperm subpopulations in each species and among species. Values are mean±SEM. *Significantly different vs. corresponding control (p<0.05). Different letters show significant difference (p<0.05) |
DISCUSSION
Nitric oxide (NO) is an important physiological messenger and effector molecule in many biological systems, including immunological, neuronal, reproductive and cardiovascular tissues (Bredt and Snyder, 1994). Due to its involvement in these diverse systems, interest in measuring NO in biological tissues and fluids remains strong. One means to investigate nitric oxide formation is to measure nitrite (NO2¯), which is one of two primary, stable and nonvolatile breakdown products of No. In this study for evaluation of basal No production in different motile sperm populations, we measured nitrite in culture media after 24 h incubation of sperm (fractionated by Percoll gradient).
There are two schools of thought as to the role of NO in the maintenance of sperm motility. Detrimental inhibitory effects have been reported by Rosselli et al. (1995) and Weinberg et al. (1995) when millimolar concentrations of exogenous NO donors were added to sperm samples. Donnelly et al. (1997) showed that basal release of NO by spermatozoa from normozoospermic samples tended to be greater than that from asthenozoospermic samples, suggesting a physiological and beneficial role for endogenous NO in the preservation of sperm motility. This is in accordance with the previous observations that normozoospermic spermatozoa express more NOS and generate more nitrite than asthenozoospermic spermatozoa (Lewis et al., 1996). These studies are not agreement with present study, because we found that nitrite production in low density sperm subpopulation with low quality and motility, didn’t show significant difference vs. high density sperm subpopulation with high quality and motility. Donnelly et al. (1997) has also demonstrated that there was little difference in basal NO production between samples with a good (>14% normal) or poor (<14% normal) morphology. This study suggested that poor quality semen is not associated with elevated basal NO production and that the effects of Reactive Oxygen Species (ROS) on sperm membranes, which depress sperm function (Balercia et al., 2004; Iwasaki and Gagnon, 1992) are not mediated by basal NO per se. these data confirm our results. O’Brayan et al. (1998) also showed that there was significant coloration between aberrant patterns of human sperm eNOS expression and decreased sperm motility. These data were inconsistence with Lewis et al. (1996), Donnelly et al. (1997) and present results.
Taken to gather, the present study showed that Basal No is produced by human, bull and ram spermatozoa, but in the same sample, there were no significant difference between high density (with higher percentage of motility) sperm subpopulation and low density (with lower percentage of motility) sperm subpopulation obtained by Percoll gradient centrifugation. Therefore, it seems that spermsatozoa with different motility in the one sperm population have the same production of NO in physiological condition that is different from pathological condition (e.g., in the subfertile men) studied by others researchers.
ACKNOWLEDGMENTS
We are grateful to Dr. Teshfam for their excellent help in performing this study. This study was supported by Islamic Azad University (Karaj branch) Research Grant.
REFERENCES
- Balercia, G., S. Moretti, A. Vignini, M. Magagnini and F. Mantero et al., 2004. Role of nitric oxide concentrations on human sperm motility. J. Androl., 25: 245-249.
Direct Link - Bredt, D.S. and S.H. Snyder, 1994. Nitric oxide: A physiologic messenger molecule. Ann. Rev. Biochem., 63: 175-195.
CrossRefDirect Link - Burnett, A.L., 2006. Nitric oxide in the penis-science and therapeutic implications from erectile dysfunction to priapism. J. Sex. Med., 3: 578-582.
Direct Link - Coleman, J.W., 2001. Nitric oxide in immnity and inflammation. Int. Immunopharm., 1: 1397-1406.
Direct Link - De Lamirande, E. and C. Gagnon, 1991. Quantitative assessment of the serum-induced stimulation of human sperm motility. Int. J. Androl., 14: 11-22.
Direct Link - Donnelly, E.T., S.E.M. Lewis, W. Thompson and U. Chakravarthy, 1997. Sperm nitric oxide and motility: The effects of nitric oxide synthase stimulation and inhibition. Mol. Hum. Reprod., 3: 755-762.
Direct Link - Herrero, M.B., S.P. Martinez, J.M. Viggiano, J.M. Polak and M.F. de Gimeno, 1996. Localization by indirect immunofluorescence of nitric oxide synthase in mouse and human spermatozoa. Reprod. Fertil. Dev., 8: 931-934.
Direct Link - Iwasaki, A. and C. Gagnon, 1992. Formation of reactive oxygen species in spermatozoa of infertile patients. Fertil. Steril., 57: 409-416.
PubMedDirect Link - Lewis, S.E.M., E.T. Donnelly, E.S.L. Sterling, M.S. Kennedy, W. Thompson and U. Chakravarthy, 1996. Nitric oxide synthase and nitrite production in human spermatozoa: Evidence that endogenous nitric oxide is beneficial to sperm motility. Mol. Hum. Reprod., 2: 873-878.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - McCann, S.M., C. Mastronardi, A. Walczewska, S. Karanth, V. Rettori and W.H. Yu, 1999. The role of nitric oxide in reproduction. Brazil. J. Med. Biol. Res., 32: 1367-1379.
Direct Link - Meiser, H. and R. Schulz, 2003. Detection and localization of two constitutive NOS isoforms in bull spermatozoa. Anat. Histol. Embryol., 32: 321-325.
Direct Link - Michel, T. and O. Feron, 1997. Nitric oxide synthases: Which, where, how and why? J. Clin. Invest., 9: 2146-2152.
Direct Link - Pacher, P., J.S. Beckman and L. Liaudet, 2007. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev., 87: 315-424.
CrossRefPubMedDirect Link - Rosselli, M., R. Dubey and B. Imthurn, 1995. Effect of nitric oxide on human spermatozoa: Evidence that nitric oxide decreases sperm motility and induces sperm toxicity. Hum. Reprod., 10: 1786-1790.
Direct Link - Telfer, J.F., F. Lyall, J.E. Norman and I.T. Cameron, 1995. Identification of nitric oxide synthase in human uterus. Hum. Reprod., 10: 19-23.
Direct Link - Weinberg, J.B., E. Doty, J. Bavaventena and J.F. Haney, 1995. Nitric oxide inhibition of sperm motility. Fertil. Steril., 64: 408-413.
Direct Link - Burnett, A.L., D.D. Ricker and S.L. Chamness, 1995. Localization of nitric oxide synthase in the reproductive organs of male rat. Biol. Reprod., 52: 1-7.
Direct Link