
Ch. Muhammad Ishtiaq
Department of Chinese Medicine Science and Engineering,
College of Pharmaceutical Sciences, Institute of Bioinformatics,
Zhejiang University, No. 38, Zhe da Road Hangzhou, 310027, Peoples Republic of China
Q. He
Department of Chinese Medicine Science and Engineering,
College of Pharmaceutical Sciences, Institute of Bioinformatics,
Zhejiang University, No. 38, Zhe da Road Hangzhou, 310027, Peoples Republic of China
S. Feng
Department of Chinese Medicine Science and Engineering,
College of Pharmaceutical Sciences, Institute of Bioinformatics,
Zhejiang University, No. 38, Zhe da Road Hangzhou, 310027, Peoples Republic of China
Yi Wang
Department of Chinese Medicine Science and Engineering,
College of Pharmaceutical Sciences, Institute of Bioinformatics,
Zhejiang University, No. 38, Zhe da Road Hangzhou, 310027, Peoples Republic of China
P. G. Xiao
Institute of Medicinal Plant Development,
Chinese Academy of Medical Sciences, Beijing, 100094, Peoples Republic China
YiYu Cheng
Department of Chinese Medicine Science and Engineering,
College of Pharmaceutical Sciences, Institute of Bioinformatics,
Zhejiang University, No. 38, Zhe da Road Hangzhou, 310027, Peoples Republic of China
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 12 | Page No.: 2066-2072







ABSTRACT
Distribution patterns of chemical compounds in plants have been used for biosystematic and phylogenetic studies. Saponin profile of twelve major taxa of Clematis genus, belonging to sections, Rectae, Clematis, Meclatis, Tubulosae and Viorna were analyzed by HPLC coupled with diode array detector and ESI-MS. The chemodiversity profile of saponins has unambiguously delimited the taxa of Clematis at subgenus, section and subsection level. The distribution of saponins in Clematis genus provides useful taxonomic markers and results are presented in phenograms. The compound Huzhangoside D was common and the most abundant in analyzed species of the genus. The morphological analysis was also conducted of the same taxa and presented as cluster tree. The distribution and chemotaxonomic importance of saponins profile within the genus is discussed.
PDF Abstract XML References Citation
How to cite this article
Ch. Muhammad Ishtiaq, Q. He, S. Feng, Yi Wang, P. G. Xiao and YiYu Cheng, 2007. Chemodiversity of Saponins and their Taxonomic Importance in Clematis Genus (Ranunculaceae). Pakistan Journal of Biological Sciences, 10: 2066-2072.
DOI: 10.3923/pjbs.2007.2066.2072
URL: https://scialert.net/abstract/?doi=pjbs.2007.2066.2072
DOI: 10.3923/pjbs.2007.2066.2072
URL: https://scialert.net/abstract/?doi=pjbs.2007.2066.2072
INTRODUCTION
Clematis L. is the second largest genus of Ranunculaceae with more than 300 species worldwide, 147 (93 endemic) in China (Wang, 1999). Clematis genus is medicinally very important because it has been used in Traditional Herbal Medicines (THM) to cure gonorrhoea (Armando et al., 1995). HIV syndrome (Byung et al., 2001), anti-tumor activity (Qiu et al., 1999), anti-inflammation and analgesic (Wang et al., 1998), hepatic protective (Chiu et al., 1988), anti-fungal (Zhizhi et al., 2003), rheumatism, fever, infections, edemas (Yesilada et al., 1997; Ch. Mahammad Ishtiaq et al., 2006) it is used to promote blood circulation, cure urinary tract infection, nephritis, amenorrhea and scanty of lactation (Ishtiaq et al., 2006; 2007).
Traditional classification systems primarily relying on floral and vegetative characters have been used for the taxonomic divisions of the genus (Tamura, 1966-1968). In past, morphological based taxonomic research has been carried out on various taxa of the Clematis genus (Tobe, 1974, 1980; Tarasevich and Serov, 1986; Snoeijer, 1992; Yano, 1992). Some of previous phylogenetic approaches based on floral and vegetative characters placed Clematis and Anemone in the same tribe (Tamura, 1967). Later on Hoot also included Clematis in the same clade with Anemone, depending on characteristics of achene morphology and presence of chemical compound ranunculin (Hoot, 1995). However, classification based on characters of seedling and juvenile morphology has been cited in recent decades as supporting a fundamental division in the infrageneric classification of the genus (Tamura, 1987). Recently in China, several attempts based on morphological characters have been carried out to study the phylogenetic position of Chinese Clematis (Hua and Li, 2003; Wang, 1998; 2000b; 2001; 2002; 2003; 2004a-c; Wang and Li, 2005a, b; Yang and Huang, 1992). However, some taxa of the genus; subsection Clematis and subsection Rectae and subsection Connatae and subsection Crispae are so closely related to each other that it is difficult to ascertain the systematic position of some intermediate species between the two subsections of each pair (Wang, 1998). On chemical grounds, Dannis used the flavonoid profiles for the chemotaxonomy study of subsection Viornae (section Viorna), of Clematis genus (Dannis, 1976). Saponins have been used as chemotaxonomic markers in differentiating other taxa of plants (Michael, 1993). Although, saponins have been isolated from different species of Clematis in the previous studies (Dekanosidze, 1979; Ayhan, 1970; He et al., 2001; Zhizhi et al., 2003; Bahcguna, 1989; Baoping et al., 1995, 1996; Haruhisa et al., 1995; Thapliyal and Bahuguna, 1993; Sati and Sudhir, 1992; Sati et al., 1990; Yukio, 2001; Hui et al., 2000) but hitherto no attempt has been conducted for chemotaxonomy of Clematis genus on the basis of saponin profile. We first time, describe distribution patterns of saponins and their taxonomic importance in the genus. The degree of chemodiversity and its potential significance as chemosystematic aspects are discussed and results are compared with published molecular and morphological classification data on Clematis genus.
MATERIALS AND METHODS
Chemicals and plant material: Acetonitrile, methanol, CHCl3 and n-butanol were of analytical grade (Merck, USA). HPLC grade water was prepared by Milli-pore water purification system (Millipore, MA, USA). The plant specimens were collected from Tian Mu Shan Biosphere reserve (TMSBR) and Hangzhou (HZ), Wenzhou, Hebei and Guangdong. The species were identified and their herbaria with voucher number were prepared (Table 1). The voucher specimens were deposited in College of Pharmaceutical Sciences, Department of Chinese Medicine Science, Zhejiang University Hangzhou, China.
Phytochemical analysis
Extraction: Plant material was air dried in dark at room temperature and powdered. For extraction ca. 1 g of powder was refluxed in 20 mL CHCl3 for 1 h and filtered.
The filtrate was discarded and the residue (plant material) was refluxed again in 40 mL of 50 % MeOH for 1 h. Extract was filtered and concentrated at 60°C under vacuum by Buchi rotavapor B-490. The concentrate was dissolved in 5 mL of dist. water and mixed with 7 mL of n-butanol in separator funnel. After an interval, mixture was partitioned into two layers and under layer was separated and stored as fraction I. The process was repeated once for residue left in funnel and isolated fraction was mixed with first one. The obtained fractions were concentrated and dissolved in 5 mL of MeOH and stored as stock solution at 4°C until use.
Sample preparation: The standard compounds ca. 1.5 mg were dissolved in 500 μL of methanol and stored as stock solution at 4° C until use. The stock solution of sample ca. 1.2 mL was centrifuged and used for analytical run in HPLC, for each specimen. About 200 uL standard solution was run to calibrate the analytical conditions.
Analytical HPLC with diode array and ultra violet detection (HPLC-DAD): The HPLC-UV analysis was carried out on an Agilent 1100 Series HPLC with diode array detector using a 5 μm Agilent RP column C18 (4.00 250 mm). The column temperature was maintained at 30°C. Optimum detection wavelength was 204-208 nm. A gradient binary elution: acetonitrile (A) and 0.1% aqueous formic acid (B). The mobile phase flux conditions; 0-5 min, 5-10% A; 6-18 min, 10-17% A; hold isocratic elution for 10 min; 29-60 min, 25% A; 61-75 min, 95% A; 76-80 min, 95% A, were used in the analysis. An auto-sampler system was used for sample injections (20 μL) and flow rate was 0.8 mL min-1. Minimum re-equilibrium time between two injections was 15 min and each sample was analyzed twice from the same vial.
| Table 1: | Plant sources and their geographical distribution and herbarium numbers |
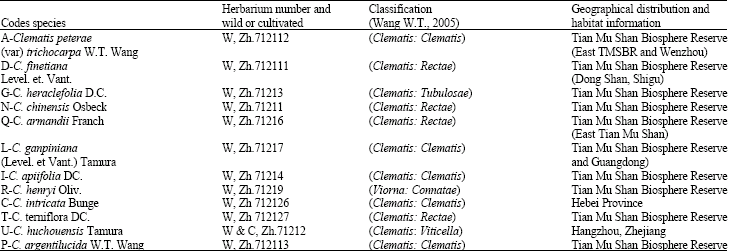 | |
Abbreviations used above: W: Wild; C: Cultivated; Zh: Zhejiang University Herbarium, species are arranged according to classification system of Wang (2005) | |
| Table 2: | HPLC retention times, UV absorption maxima and molecular weight [MH+(m/z)] |
 | |
| Table 3: | Saponin distribution in the Clematis genus using HPLC-UV and ESI-MS |
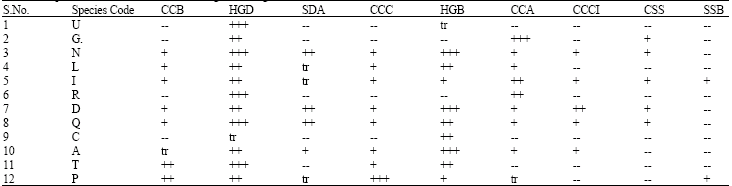 | |
Abbreviations used above: CCB: Clematichinoside B; HGB: Huzhangoside D; SDA: Seiboldianoside A; CCC: Clematichinoside; HGB: Huzhangoside B; CCA: Clematichinenoside A; CCCI: Clematichinoside Isomer; CSS: Clemastanoside; SSB: Songaroside B; +++: large concentration; ++: large to moderate concentration; +: minor concentration; tr: trace concentration; --: not detectable | |
High performance liquid chromatography coupled with mass spectrometry (HPLC-ESI-MS): HPLC-MS was performed with an 1100 Series HPLC and quadrupole ion trap mass spectrometer (ThermoFinnigan LCQ-DECAXPlus). The HPLC conditions were same as above mentioned. The mass spectra were recorded using quadrupole ion trap mass spectrometer with the sample ionized by an ESI source operated in negative mode and using vaporizer temperature 550°C, sheath and auxillary nitrogen flow pressures of 30 and -10 Ψ, respectively. Capillary temperature 350° C and capillary voltage -15° C were optimum in this analysis. The mass spectrometer was controlled by Xcalibur 1.3 software (ThermoFinnigan) and programmed to record survey scans in the range m/z 200-2000, in TIC mode. The recorded data were analyzed and identified by comparison their retention times, UV spectra and TIC patterns in MS with standards and cited literature. The peak area variations of those compounds were calculated which were common in at least two species. The qualitative and quantitative variations of different compounds in the analyzed taxa were formulated in the form of a matrix. The calculated matrix was used for constructing a Chemotypic Cluster Tree (CCT) by hierarchical cluster approach euclidian distance, average link (Classical Unweighted Pair-Group Method Using Arithmetic Averages) algorithm by the program in Matlab 6.5 (Mathwork Inc.). The CCT presented in the form of a phenogram indicates infrageneric position of different taxa of Clematis genus (Fig. 1).
RESULTS AND DISCUSSION
Identification of the saponins: For the identification of saponins in extracts of different taxa of Clematis, their HPLC Retention Times (RT), Ultraviolet Spectra (UV) and ESI mass spectra were compared with those of standards previously isolated from Clematis ganpiniana (Sun et al., 2007) or with saponins present in the genus which had been identified and cited in literature (Shao et al., 1995; Chirva et al., 1974; Shao et al., 1996; Kizu et al., 1995; Kawata et al., 1998; Song et al., 1992). The RT, UV spectra and ESI mass spectra of the peaks of the analyzed samples of Clematis are presented in the Table 2. Only five standards of saponins were available for chromatographic comparison (Huzhangoside B, Clematichinenoside C, Seiboldianoside A, Huzhangoside D, Clematichinenoside B), so that other saponins detected in the analysis could not be identified by HPLC. No standard compounds were available for the compounds 6-9, but their RT, UV spectra and MS spectra from ESI source were compared with previously identified compounds of the genus and four compounds were tentatively found to be Clemochinenoside A, Songaroside B, Clemastanoside A, Clematichinenoside C (isomer) with molecular weight 684.6, 1028.5, 1378.6 and 1499 by LC-ESI-MS.
Presence and distribution of saponins in Clematis species: This distribution pattern of saponins mirrors taxonomic relationships among the taxa and predict their ecological and morphological characteristics.
| Table 4: | Characters and character states used in the morphological-based cladistic analysis |
 | |
The distribution of saponin profile of Clematis genus is given in Table 3. Among the identified compounds; Huzhangoside D had high concentration in C. chinensis, C. henryi, C. armandii and C.terniflora and Huzhangoside B showed high quantity in C. chinensis, C. huchouensis, C. finetiana and C. peterae. Clemochinenoside A depicted high amount in C. heracleifolia and C. armandii and other compounds were found moderate to minor amounts or some times as traces in different analyzed species (Table 3). To see whether there is any correlation between distribution of saponins and taxonomy of Clematis genus based on morphological characters, the analyzed species were morphologically studied. Out of fifty morphological characters surveyed, 27 characters were selected for phylogenetic study because of their low infraspecific variation, presence in the most of taxa, ability to be scored unequivocally and phylogenetic informativeness (Table 4). These phenetic characters were used to generate a morphological cluster tree (MCT) which was almost similar to the revised classification system of Wang (Fig. 1).
On the basis of presence or absence of saponin compounds and their peak area variation in different taxa, a matrix was formulated. A chemical cluster tree (CCT) was constructed from this matrix data using euclidian distance, average link (Classical Unweighted Pair-Group Method Using Arithmetic Averages) algorithm by Matlab software (Fig. 2). According to chemical profile of saponins, analyzed taxa of the genus are divided into three main clades presented in CCT (Fig. 2). First group (clade I) of CCT includes species; Clematis finetiana (D), C. armandii (Q), C. chinensis (N) and C. terniflora (T) and these species belong to subsection Rectae, albeit on morphological bases they are very closely related yet well isolated in this chemical analysis (Fig. 2). The clade I species share all five standard compounds and are fairly aggregated as one clade branch of CCT but still they are well separated at species level due to quantity variation.
The clade II (subsection Clematis) consists of C. apiifolia (L), C. argentilucida (I), C. ganpiniana (P) and C. peterae (A) species which are morphologically very similar but fairly resolved in chemotaxonomic analysis. The clade II (L, I, P, A) species share compounds CCB, HGD, CCC and HGB with high to minor concentration and are re-grouped as one aggregate in CCT (Fig. 2), however individual species are well separated due to quantitative variation in concentration of these common compounds in different taxa although very closely allied on morphological basis. Third aggregate (clade III) contains those species which belongs to different sections; C. heracleifolia (sect. Tubulosae), C. huchouensis (sect. Viticella), C. intricata (sect. Meclatis) and C. henryi (sect. Connatae) are fairly separated from each other as well as from other two most similar clades (clade I and II).
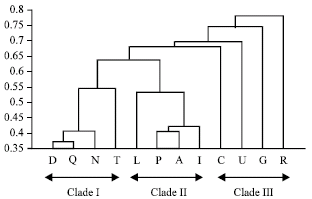 | |
| Fig. 1: | Phenogram showing affinity relationships among taxa of Clematis genus based on morphological characters, as determined by Euclidian distance, average link (Classical Unweighted Pair-Group Method Using Arithmetic Averages) algorithm. The alphabetical names are same as presented in Table 1 |
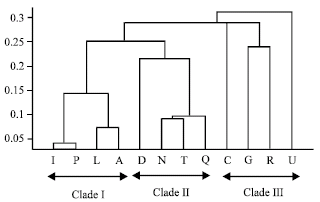 | |
| Fig. 2: | Phenogram showing affinity relationships among taxa of Clematis genus based on chemotypic characters, as determined by Euclidian distance, average link (Classical Unweighted Pair-Group Method Using Arithmetic Averages) algorithm. The alphabetical names are same as presented in Table 1 |
One discrepancy was observed that species R (C. henryi) predicted close affinity with species G (C. heracleifolia) and appeared as sister branch in CCT but on morphological basis it appeared as separate, outer most individual line. Another difference between two approaches is that in CCT species U (C. huchouensis) occupied the most distant position predicting more intra-clade and inter-clade differences while in MCT it has more affinity with them. This may be due to that out of many taxa we used only few species representing the major taxa of Clematis genus or due to less chemical data incorporated in CCT, as we were able to use only few compounds in this analysis, due to lack of standard compounds. Among these, C. huchouensis (U) sample possessed these saponin compounds (HGD, SDA, CCC, HGB) and appear as one line in CCT, C. heracleifolia (G) consists of compounds (HGD, CCA, CSS) and C. intricata © has compounds (HGB, HGD) but latter one (HGD) in trace. The species C. henryi (R) belongs to section Connatae (subgenus Viorna) has compounds (HGD, CCA) common with other species heracleifolia (G) and appears sister clade branch with species G in third clade of CCT. The chemical data predicts that they may be genetically more similar to each other than other members of clade III. The compound 3 (SDA) seems to be mostly restricted with moderate quantity to clade I (D, N, Q, T) making it special chemical marker for its identification. However distinctive characteristics of subsection Clematis are presence of compounds HGD, SDA, CCC in large to minor quantity. Furthermore, compounds CCB, SDA, CCC, HGB, CCCI, CSS and SSB can be helpful in demarcating taxa boundaries at subgenus level in Clematis genus. The results of CCT congruently favour MCT and previously reported traditional and molecular based classification systems of Clematis genus (Wang, 2005: Jonathan, 2004). But some substantial differences among species G and R and other group III taxa are still bottleneck. So, further detailed and comprehensive phytochemical research is required in order to designate their distinctive taxonomic position. Moreover, future chemotaxonomic analysis including much more taxa of Clematis genus is inevitable for better understanding of profound taxonomical and phylogenetic status of various taxa of the Clematis genus.
CONCLUSIONS
The chemotypic profile of saponins well represents and demarcates infrageneric relationships of Clematis genus taxa in conjunction with morphological evidences (Fig. 1 and 2). The closely nested species of Clematis are fairly delimited by saponin distribution patterns on basis of qualitative and quantitative variations. The analyzed taxa of Clematis are differentiated into well separated clades and ubiquitously distinguish the infraspecific relationships among the subsect. Clematis and Rectae (sect. Clematis) and other analyzed taxa. But still show some substantial differences at proximal points (clade III) exist from classical approach on the genus. Hence, detailed morphological and chemotaxonomic analysis throughout the whole range of distribution of Clematis taxa may be helpful to study the comprehensive phylogenetic and taxonomic position of this large and complex genus of Ranunculaceae.
ACKNOWLEDGMENTS
The authors wish thanks to staff members of the Tian Mu Shan Biosphere Reserve, who helped in the collection of plants specimens. Special regards to Prof. Chen J.H., who did a lot of help in field trips and identification of specimens. Thanks to others members of laboratory who helped directly or indirectly in this research.
REFERENCES
- Armando, C., M. Herlina, M. Emilia, C. Ericka and E.S. Blanka et al., 1995. Antigonorrhoeal activity of plants used in Guatemala for treatment of sexually transmitted diseases. J. Ethnopharmacol., 48: 85-88.
Direct Link - Baoping, S., Q. Guowei, X. Rensheng, W. Houming and M. Kan, 1995. Triterpenoid saponins from Clematis chinensis. Phytochemistry, 38: 1473-1479.
Direct Link - Baoping, S., Q. Guowei, X. Rensheng, W. Houming and M. Kan, 1996. Saponins from Clematis chinensis. Phytochemistry, 42: 821-825.
Direct Link - Du, Z., N. Zhu, N. Ze-Ren-Wang-Mu and Y. Shen, 2003. Two new antifungal saponins from the Tibetan herbal medicine Clematis tangutica. Planta Medica, 69: 547-551.
Direct Link - Haruhisa, K., S. Hideki and T. Tsuoshi, 1995. Studies on constituents of Clematis species VI. Clematis stans Sieb et Zucc. Chem. Pharm. Bull, 43: 2187-2194.
Direct Link - He, M. and J.H. Zhang and C.Q. Hu, 2001. Studies on chemical components of Clematis chinensis. Acta Pharma. Sin., 36: 278-280.
Direct Link - Hoot, S.B., 1995. Phylogeny of Ranunculaceae based on preliminary atpB, rbcL and 18S nuclear ribosomal DNA sequence data. Plant Syst. Evol., 9: 241-251.
Direct Link - Hua, S.J. and L.Q. Li, 2003. Leaf epidermal feature in Clematis (Ranunculaceae) with reference to its systematic significance. Acta Bot. Sin., 45: 257-268.
Direct Link - Hui, M.Z., X.C. Zhang, T. Xuan, X.C. Yu and Z.C. Yao, 2001. Triterpenoid Saponins from Clematis tangutica. Planta Med., 67: 484-488.
Direct Link - Kawata, Y., H. Kizu and T. Tomimori, 1998. Studies on the constituents of Clematis species. VII. Triterpenoid saponins from the roots of Clematis terniflora DC. var. robusta Tamura. Chem. Pharm. Bull., 46: 1891-1900.
Direct Link - Kizu, H., H. Shimana and T. Tomimori, 1995. Studies on the constituents of Clematis Species VI. The constituents of Clematis stans Sieb. Chem. Pharm. Bull., 43: 2187-2194.
Direct Link - Michael, F., J. Fay, Philip and J. Dale, 1993. Condensed tannins in Trifolium species and their significance for taxonomy and plant breeding. Genet. Res. Crop Evol., 40: 7-13.
Direct Link - Chaudhary, M.I., Q. He, Y.Y. Cheng and P.G. Xiao, 2006. Ethnobotany of medicinal plants from tian mu shan biosphere reserve, zhejiang-province, China. Asian J. Plant Sci., 5: 646-653.
CrossRefDirect Link - Muhammad, I. Ch., Q. He, P.G. Xiao and Y.Y. Cheng, 2007. Clematis huchouensis TAMURA: A traditional Chinese herbal medicine and its quality control using a high performance liquid chromatography technique. Biol. Pharm. Bull., 30: 165-168.
Direct Link - Sati, O.P., S.K. Uniyal, S. Bahcguna and T. Kikuchi, 1990. Clematosides. A triterpenoid saponin from the roots of Clematis grata. Phytochemistry, 29: 3676-3678.
Direct Link - Sati, O.P. and K.U. Sudhir, 1992. Triterpenoid saponins from roots of Clematis grata. Phytochemistry, 31: 1427-1428.
Direct Link - Baoping, S., Q. Guowei, X. Rensheng, W. Houming and M. Kan, 1995. Triterpenoid saponins from Clematis chinensis. Phytochemistry, 38: 1473-1479.
Direct Link - Wang, W.T., 2000. Notes on the genus Clematis (Ranunculaceae) (III). Acta Phytotax. Sin., 38: 497-514.
Direct Link - Wang, W.T., 2002. A revision of clematis sect. Cheiropsis (Ranunculaceae). Acta Phytotax. Sin., 40: 193-241.
Direct Link - Wang, W.T., 2004. A revision of Clematis sect. Aspidanthera s.l. (Ranunculaceae). Acta Phytotax. Sin., 42: 97-135.
Direct Link - Wang, W.T., 2004. A revision of Clematis sect. Brachiatae (Ranunculaceae). Acta Phytotax. Sin., 42: 289-332.
Direct Link - Wang, W.T., 2004. A revision of Clematis sect. Pseudanemone (Ranunculaceae). Acta Phytotax. Sin., 42: 385-418.
Direct Link - Yesilada, E., O. Ustun, E. Sezik, Y. Takaishi, Y. Ono and G. Honda, 1997. Inhibitory effects of Turkish folk remedies on inflammatory cytokines: Interleukin-1α, interleukin-1β and tumor necrosis factor α. J. Ethnopharmacol., 58: 59-73.
CrossRef