
Parviz Malekzadeh
Department of Biology, Faculty of Science, Urmia University, Urmia, Iran
Jalil Khara
Department of Biology, Faculty of Science, Urmia University, Urmia, Iran
Shadi Farshian
Department of Biology, Faculty of Science, Urmia University, Urmia, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 12 | Page No.: 2008-2013







ABSTRACT
Soil microorganisms have been shown to possess several mechanisms capable of altering metal bioavailability for uptake into roots. In addition, root mycorrhizal associations have been shown to affect the rate of metal uptake. There is evidence that exposure of plants to excess concentrations of heavy metals such as Cu results in oxidative injury. In this study, effect of arbuscular mycorrhizal fungus Glomus etunicatum on tolerance of Cu toxicity in tomato plants was studied. In order to prepare seedling medium, we used washed and sterilized sand and agricultural soil. Tomato seeds were surface sterilized and planted in two pots. One filled just with sterilized sand (for non-mycorrhizal treatments) and the other filled with sterilized sand mixed with G. intraradices mycorrhizal inoculum. We were certain about complete colonization after 4 weeks, so we transferred three seedlings to each main pot. Plants grew in growth chamber for nine weeks. During growth period plants received modified Hoagland’ s solution (with half P content) with Cu concentration of 0, 1.5, 3.5, 5.5, 7.5 mM CuSO4 in triplicates. Antioxidant enzymes activity, Ascorbate Peroxidase (APX) and Guaiacol Peroxidase (GPX) and Root Length Colonization (RLC) percentage in mycorrhizal and non-mycorrhizal plants were measured. APX activity in mycorrhizal shoots increased but there was no significant correspondent increase in roots of these plants. GPX activity in mycorrhizal roots increased but there was no significant correspondent increase in shoots of these plants. Activity of this enzyme in roots and shoots of mycorrhizal plantshigher than non-mycorrhizal plants. Estimation of root length colonizatinon by gridline intersect method, increase in Cu concentration, colonization percentage decreased significantly. The data show the possible role of mycorrhiza in plant protection against Cu toxicity.
PDF Abstract XML References Citation
How to cite this article
Parviz Malekzadeh, Jalil Khara and Shadi Farshian, 2007. Copper Toxicity Influence on Antioxidant Enzymes Activity in Tomato Plants and Role of Arbuscular Mycorrhizal Fungus Glomus etunicatum in the Tolerance of Toxicity. Pakistan Journal of Biological Sciences, 10: 2008-2013.
DOI: 10.3923/pjbs.2007.2008.2013
URL: https://scialert.net/abstract/?doi=pjbs.2007.2008.2013
DOI: 10.3923/pjbs.2007.2008.2013
URL: https://scialert.net/abstract/?doi=pjbs.2007.2008.2013
INTRODUCTION
Copper (an essential micronutrient) is frequently related to soil pollution and is potentially toxic to living organisms. Copper (Cu) is necessary for plant growth in low concentrations, a structural part of enzymes and is uptaken as divalent cation (Cu2+) or Cu chelate. Cu, however, is often present in high concentrations that are toxic enough to biota.
When higher plants are subjected to environmental stresses, such as water stress and salinity, a variety of Reactive Oxygen Species (ROS) such as superoxide anion radical (O2_), hydrogen peroxide (H2O2), hydroxyl radicals (OH¯) and singlet oxygen (1O2) are induced (Elstner, 1982; Jung, 2004). As a consequence, higher plants induce efficient antioxidant systems to protect them against oxidative injury (Asada, 1999). The antioxidant systems consist of antioxidant enzymes including superoxide dismustase (SOD), Guaiacol Peroxidase (G-POD), catalase (CAT), Glutathione Reductase (GR), Ascorbate Peroxidase (APX) and non-enzymatic antioxidants including ascorbate (ASC) and glutathione (GSH), which are designed to minimize the concentrations of O2d_ and H2O2.
Arbuscular Mycorrhiza (AM) is one of the most widespread Mycorrhizal associations between soil microorganisms and higher plants (Malekzadeh et al., 2007a). Ninety percent of the earth’ s land plant species form symbiotic associations with Arbuscular Mycorrhizal Fungi (AMF) (Gadkar et al., 2001). Providinga direct physical link between soil and plant roots, the Arbuscular Mycorrhiza (Am) fungi are important rhizospheric microorganisms. They can increase plant uptake of nutrients especially relatively immobile elements such as P, Zn and Cu (Ryan and Angus, 2003) and consequently, they increase root and shoot biomass and improve plant growth. It has been indicated that Am fungi can colonize plant roots in metal contaminated soil (Vogel-Mikus et al., 2005), while their effects on metal uptake by plants are conflicting. In slightly metal contaminated soil, most studies show that Am fungi increased shoot uptake of metals (Weissenhorn et al., 1995), while in severely contaminated soil, Am fungi could reduce shoot metal concentration and protect plants against harmful effects of metals (Li and Christie, 2001; Malcova et al., 2003).
As a result of this symbiotic association between AM fungi and host plants, P content also has an effect on physiological parameters in plants (Johnson, 1984; Paradi et al., 2003). One of the physiological parameters is the increase in photosynthesis.
AMF-colonized plants are generally more resistant to stresses caused by drought, salt, heavy metals or attack by pathogens. These positive effects of the fungi on the growth of plants often result from an improved nutrient supply and can partly be due to complex and not easily resolved interactions between the symbiotic partners. In contrast, AMF-colonized roots of plants from soils severely polluted by high concentrations of heavy metals show lower amounts of heavy metals than non-colonized plants (Schuepp et al., 1987; El-Kherbawy et al., 1989; Weissenhorn et al., 1995; Weinstein et al., 1995; Kaldorf et al., 1999; but see also opposite data by (Gildon and Tinker, 1983; Killham and Firestone. 1983), (Rufyikiri et al., 2000; Ottet et al., 2002; Rabie, 2005; Vogel-Mikus et al., 2005).
The present study was undertaken to thoroughly evaluate reactive oxygen metabolism, including ROS, antioxidant enzymes and non-enzymatic antioxidants, in leaves and roots of tomato plant inoculated with G. etunicatum under Cu toxicity conditions.
However, information about activities of antioxidant enzymes in AM and non-AM plants is scarce.
MATERIALS AND METHODS
Seeds of tomato (Lycopersycium esculentum Mill) were germinated in a moist mix of soil and sand in polystyrene trays. There 28 day old seedlings, uniform in size, were transplanted into 30 plastic pots filled with sterilized sand. Half of the pots received the AMF Glomus etunicatum and Trappe by placing 30 g (moist weight) of inoculums is soil below the tomato seedling prior to planting. The AMF inoculum (consist of soil and root fragments and spores) (This inoculum was prepared in trap culture of G. etunicatum in the greenhouse) was placed directly adjacent to each seedling root to facilitate fungal colonization of plant roots. Control treatments received no AMF inoculum.
The plants were grown in a greenhouse in Urmia university under natural photoperiods (28/20°C day/night 6000/10000 lux light in tensity) for 13 weeks during which only distilled water was applied. In addition, twice a week, each pot was supplied with 100 mL of a nutrient solution containing: five Cu concentration (0, 1.5, 3.5, 5.5, 7.5 Mm CuSO4) added to Hogland nutrient solution (with half P concentration).
The symbiotic fungal partner, Glomus etunicatum, was produced in a soil:and (1/1, v/v) mixture using maize as the host plant. Inoculum of Glomus etunicatum (30 g), consisting of spores, external mycelium and AMF colonized roots, was laid around the seed. The same amount of sterilized inoculum was laid into the control pots. The roots were cleared and stained by using the methods by Philips and Hayman (1970). The percentage of mycorrhizal colonization (F % ) was estimated by the grid line intersect method (Giovanetti and Mosse, 1980). At the end of the experiment, plants were harvested 10 weeks after seed sowing. Plant shoots was separated, dried (70°C 48 h).
For extraction of antioxidative enzymes, roots were homogenized with 0.1 M sodium phosphate buffer (pH 6.8) in a chilled pestle and mortar. The homogenate was centrifuged at 12,000 g for 20 min and the resulting supernatant was used for determination of enzyme activity. The whole extraction procedure was carried out at 4°C. The decrease in the concentration of ascorbate was recorded at 290 nm. The enzyme activity was calculated from the initial rate of the reaction using the extinction coefficient of ascorbate (2.8 mM_1 cm_1 at 290 nm).
The whole extraction procedure was carried out at 4°C. APOD and GR were assayed as described previously (Chang and Kao, 1998).
RESULTS
Figure 1 shows the effect of Cu treatment on the activity of APX in the shoots of mycorrhizal and non-mycorrhizal tomato plants. APX activity in mycorrhizal shoots increased significantly (p<0.05) by increase in Cu concentration, but there was no significant correspondent increase in roots of these plants (Fig. 2). Activity of this enzyme in non-mycorrhizal plant increased significantly (p<0.05), but Activity of this enzyme in roots and shoots of mycorrhizal plants higher than non-mycorrhizal plants.
Figure 3 shows, GPX activity in mycorrhizal roots increased significantly (p<0.05) by increase in Cu concentration, but there was no significant correspondent increase in roots of these plants (Fig. 4). Activity of this enzyme in non-mycorrhizal plant increased significantly (p<0.05), but Activity of this enzyme in roots and shoots of mycorrhizal plants higher than non-mycorrhizal plants.
Estimation of root length colonizatinon by gridline intersect method, increase in Cu concentration, colonization percentage decreased significantly. The data show the possible role of mycorrhiza in plant protection against Cu toxicity.
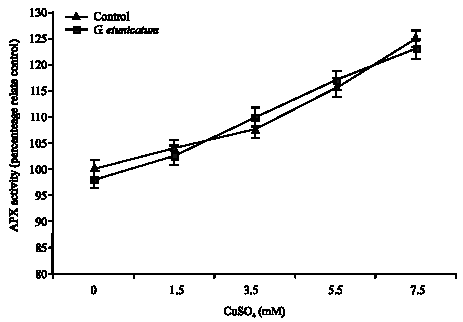 | |
| Fig. 1: | The effects of Cu treatment on the activity of APX in the shoots of mycorrhizal and non-mycorrhizal tomato plants treated with CuSO4 |
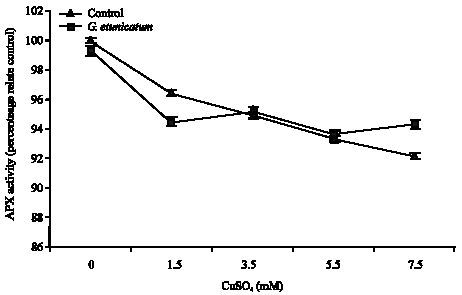 | |
| Fig. 2: | The effects of Cu treatment on the activity of APX in the roots of mycorrhizal and non-mycorrhizal tomato plants treated with CuSO4 |
Figure 5 showed that the development of Am symbiosis varied between Cu concentration. Root colonization of tomato plants was significantly reduced and negatively correlated by the presence of heavy metals in nutrient solution, it was decreased from 57% in control to 34% in the 7.5 mM concentration.
The data show the possible role of mycorrhiza in plant protection against Cu toxicity.
DISCUSSION
The protective mechanisms adapted by plants to scavenge free radicals and peroxides include several antioxidative enzymes such as SOD, APX, GR, CAT and GPX. The antioxidative enzymes are important components in preventing the oxidative stress in plants as is based on the fact that the activity of one or more of these enzymes is generally increased in plants when exposed to stressful conditions (Malekzadeh et al., 2007b).
Figure 1-4 indicates antioxidant enzyme (APX and GPX) activities of Am tomato plants showed significant higher values in polluted soil than in non-polluted one. Based on these results, the mechanisms related to physiological interactions between the Am fungus and tomato plants involve increased protein synthesis as well as induction of antioxidant enzymes to avoid heavy metal-mediated oxidative stress.
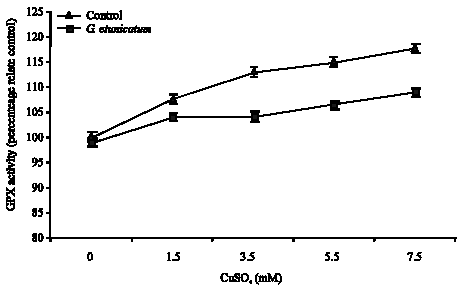 | |
| Fig. 3: | The effects of Cu treatment on the activity of GPX in the shoots of mycorrhizal and non-mycorrhizal tomato plants treated with CuSO4 |
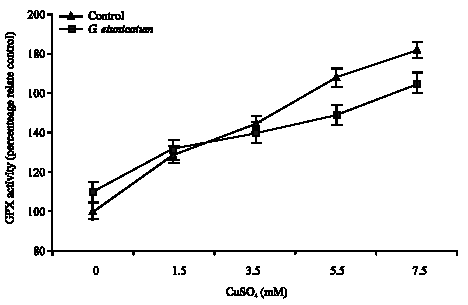 | |
| Fig. 4: | The effects of Cu treatment on the activity of GPX in the roots of mycorrhizal and non-mycorrhizal tomato plants treated with CuSO4 |
If so, the author attributed the reduced heavy metal toxicity effects in Am tomato plants to antioxidative protection through detoxification of heavy metals, chelation through metal-binding proteins (peptides) and dilution through increased plant growth induced by Am fungi. This finding supports results from some dissipated studies reporting that the role of antioxidative enzyme system as well as improved growth as possible mechanisms for plant protection against high accumulated toxic heavy metals in the shoots (Ott et al., 2002; Burleigh et al., 2003; Tong et al., 2004).
This finding supports results from numerous studies reporting that Am fungi often protect plants against high accumulation of toxic elements in the shoots, as it was reported for Al (Rufyikiri et al., 2000), Cd (Yu et al., 2004), Zn (Burleigh et al., 2003), Pb (Malcova et al., 2003), U (Rufyikiri et al., 2004), Cu (Gonzalez-Chavez et al., 2002) and As (Fitz and Wenzel, 2002).
Estimation of root length colonizatinon by gridline intersect method, resulted in 50% decrease between controls (0 mM) and 7.5 mM and showed that by increase in Cu concentration, colonization percentage decreased significantly.
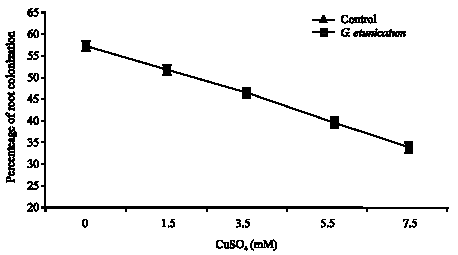 | |
| Fig. 5: | The effects of Cu treatment on the percentage of root colonization with Glomus etunicatum in tomato plants |
This finding is in line with results of Leyval et al. (1995), Weinstein et al. (1995), Hildebrandt et al. (1999) and Vogel-Mikus et al. (2005) who reported that sensitivity of Am symbionts to heavy metal contaminated soil expressed as a reduction in spore germination, hyphal growth or root colonization, had been proved previously in number of studies. Another interesting result in Fig. 5 was that the presence of Am fungi inoculations would increase the metal tolerance index of tomato plants compared with non-Am plants that grew in heavy metal polluted soil. This result emphasizes that Am fungi could be potentially effective in protecting plants exposed to high levels of heavy metal. The Am fungi ability to alleviate heavy metal stress of plants grown in heavy metal contaminated soil was previously proved by Rufyikiri et al. (2000), Hildebrandt et al. (2002) and Burleigh et al. (2003).
These results overwhelmingly indicate that the behavior of Am fungi in the protection of plants from heavy metal toxicity might differ according to host plant and metal concentration which can not generalized. Therefore additional researches are needed to explore the behavior of Am fungi in various plant species and families for plant protection in heavy metal polluted soil. Nevertheless, the beneficial effects of the Am fungus, observed in this study, arouse an interest in considering the role of Am fungi in plant-based strategies of remediation of highly heavy metal contaminated soils.
ACKNOWLEDGMENTS
The authors are grateful to Shayan Farshian, Sanam, Fradieh and Zeinab Malekzadeh for commenting on the English.
REFERENCES
- Asada, K., 1999. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol., 50: 601-639.
CrossRefPubMedDirect Link - Burleigh, S.H., B.K. Kristensen and I.E. Bechmann, 2003. A plasma membrane zinc transporter from Medicago truncatula is upregulated in roots by Zn fertilization, yet down-regulated by arbuscular mycorrhizal colonization. Plant Mol. Biol., 52: 1077-1088.
Direct Link - Chang, C.J. and C.H. Kao, 1998. H2O2 metabolism during senescence of rice leaves: Changes in enzyme activities in light and darkness. Plant Growth Regul., 25: 11-15.
Direct Link - El-Kherbawy,M., J.S. Angle, A. Heggo and R.L. Chaney, 1989. Soil pH, rhizobia and vesicular-arbuscular mycorrhizae inoculation effects on growth and heavy metal uptake of alfalfa (Medicago sativa L.). Biol. Fertil. Soils, 8: 61-65.
CrossRefDirect Link - Elstner, E.F., 1982. Oxygen activation and oxygen toxicity. Ann. Rev. Plant Physiol., 33: 73-96.
Direct Link - Fitz, W.J. and W.W. Wenzel, 2002. Arsenic transformations in the soil-rhizosphere-plant system: Fundamentals and potential application to phytoremediation. J. Biotechnol., 99: 259-278.
CrossRefPubMedDirect Link - Gadkar, V., R. David-Schwartz, T. Kunik and Y. Kapulnik, 2001. Arbuscular mycorrhizal fungi colonization factors involved in host recognition. Plant Physiol., 127: 1493-1499.
Direct Link - Gildon, A. and P.B. Tinker, 1983. Interactions of vesicular-arbuscular mycorrhizal infections and heavy metals in plants. New Phytol., 95: 263-268.
CrossRef - Giovannetti, M. and B. Mosse, 1980. An evaluation of techniques to measure vesicular-arbuscular infection in roots. New Phytol., 84: 489-500.
CrossRefDirect Link - Gonzalez-Chavez, C.D., J. D'Haen, J. Vangronsveld and J.C. Dodd, 2002. Copper sorption and accumulation by the extraradical mycelium of different Glomus spp. (arbuscular mycorrhizal fungi) isolated from the same polluted soil. Plant Soil, 240: 287-297.
CrossRefDirect Link - Hildebrandt, U., M. Kaldorf and H. Bothe, 1999. The zinc violet and its colonization by arbuscular mycorrhizal fungi. J. Plant Physiol., 154: 709-717.
Direct Link - Hildebrandt, U., E. Schmelzer and H. Bothe, 2002. Expression of nitrate transporter genes in tomato colonized by an arbuscular mycorrhizal fungus. Physiol. Plant., 115: 125-136.
Direct Link - Jung, S., 2004. Variation in antioxidant metabolism of young and mature leaves of Arabidopsis thaliana subjected to drought. Plant Sci., 166: 459-466.
Direct Link - Kaldorf, M., A.J. Kuhn, W.H. Schroder, U. Hildebrandt and H. Bothe, 1999. Selective element deposits in maize colonized by a heavy metal tolerance conferring arbuscular mycorrhizal fungus. J. Plant Physiol., 154: 718-728.
Direct Link - Leyval, C., B.R. Singh and E.J. Joner, 1995. Occurrence and infectivity of arbuscular mycorrhizal fungi in some Norwegian soils influenced by heavy metals and soil properties. Water Air Soil Pollut., 84: 203-216.
CrossRefDirect Link - Li, X.L. and P. Christie, 2001. Changes in soil solution Zn and pH and uptake of Zn by arbuscular mycorrhizal red clover in Zn contaminated soil. Chemosphere, 42: 201-207.
Direct Link - Malcova, R., M. Voslitka and M. Gryndler, 2003. Effects of inoculation with Glomus intraradices on lead uptake by Zea mays L. and Agrostis capillaris L. Applied Soil Ecol., 23: 55-67.
Direct Link - Malekzadeh, P., J. Khara and S. Farshian, 2007. Effect of arbuscular mycorrhiza (Glomus etunicatum) on some physiological growth parameters of tomato plant under copper toxicity in solution. Pak. J. Biol. Sci., 10: 1326-1330.
CrossRefPubMedDirect Link - Malekzadeh, P., J. Khara, S. Farshian, A.K. Jamal-Abad and S. Rahmatzadeh, 2007. Cadmium toxicity in maize seedlings: Changes in antioxidant enzyme activities and root growth. Pak. J. Biol. Sci., 10: 127-131.
CrossRefPubMedDirect Link - Ott, T., E. Fritz, A. Polle and A. Schutzendubel, 2002. Characterization of antioxidative systems in the ectomycorrhiza-building basidiomycete Paxillus involutus (Bartsch) Fr. and its reaction to cadmium. FEMS Microbiol. Ecol., 42: 359-366.
Direct Link - Paradi, I., Z. Bratek and F. Lang, 2003. Influence of arbuscular mycorrhiza and phosphorus supply on polymine content, growth and photosynthesis of P. lanceolata. Biol. Plant., 46: 563-569.
Direct Link - Phillips, J.M. and D.S. Hayman, 1970. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc., 55: 158-161.
CrossRefDirect Link - Rabie, G.H., 2005. Contribution of arbuscular mycorrhizal fungus to red kidney and wheat plants tolerance grown in heavy metal-polluted soil. Afr. J. Biotechnol., 4: 332-345.
Direct Link - Rufyikiri, G., S. Declerck, J.E. Dufey and B. Delvaux, 2000. Arbuscular mycorrhizal fungi might alleviate aluminium toxicity in banana plants. New Phytol., 148: 343-352.
CrossRefDirect Link - Rufyikiri, G., L. Huysmans, J. Wannijn, M. Van Hees, C. Leyval and I. Jakobsen, 2004. Arbuscular mycorrhizal fungi can decrease the uptake of uranium by subterranean clover grown at high levels of uranium in soil. Environ. Pollut., 130: 427-436.
Direct Link - Ryan, M.H. and J.F. Angus, 2003. Arbuscular mycorrhizae in wheat and field pea crops on a low P soil: Increased Zn-uptake or yield. Plant Soil, 250: 225-239.
Direct Link - Tong, Y.P., R. Kneer and Y.G. Zhu, 2004. Vacuolar compartmentalization: A second-generation approach to engineering plants for phytoremediation. Trends Plant Sci., 9: 7-9.
CrossRefDirect Link - Vogel-Mikus, K., D. Drobne and M. Regvar, 2005. Zn, Cd and Pb accumulation and arbuscular mycorrhizal colonisation of pennycress Thlaspi praecox Wulf. (Brassicaceae) from the vicinity of a lead mine and smelter in Slovenia. Environ. Pollut., 133: 233-242.
CrossRefDirect Link - Weissenhorn, I., C. Leyval, G. Belgy and J. Berthelin, 1995. Arbuscular mycorrhizal contribution to heavy metal uptake by maize (Zea mays L.) in pot culture with contaminated soil. Mycorrhiza, 5: 245-251.
CrossRefDirect Link - Weissenhorn, I., C. Leyval, G. Belgy and J. Berthelin, 1995. Arbuscular mycorrhizal contribution to heavy metal uptake by maize (Zea mays L.) in pot culture with contaminated soil. Mycorrhiza, 5: 245-251.
CrossRefDirect Link - Yu, X., J. Cheng and M.H. Wong, 2005. Earthworm-mycorrhiza interaction on Cd uptake and growth of ryegrass. Soil Biol. Biochem., 37: 195-201.
CrossRefDirect Link