
J. Sikinarum
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khan Kaen University, Khan Kaen 40002, Thailand
P. Jaisil
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khan Kaen University, Khan Kaen 40002, Thailand
S. Jogloy
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khan Kaen University, Khan Kaen 40002, Thailand
B. Toomsan
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khan Kaen University, Khan Kaen 40002, Thailand
T. Kesmala
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khan Kaen University, Khan Kaen 40002, Thailand
A. Patanothai
Department of Plant Science and Agricultural Resources, Faculty of Agriculture, Khan Kaen University, Khan Kaen 40002, Thailand
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 12 | Page No.: 1956-1962







ABSTRACT
The objectives of this study were to estimate broad sense heritability for N2 fixation and related traits and agronomic traits and evaluate correlations among these traits. Seventy four progenies in the F5 generation previously selected in the F4 generation for high and low performance for N2 fixation and related traits were tested for N2 fixation and related traits and their agronomic performances. A non-nodulating line was also included as a reference crop for nitrogen determination. The experiment was carried out under limited soil nitrogen conditions. A randomized complete block design with four replications was used. Data were recorded for leaf color score, nodule dry weight, top dry weight, fixed nitrogen, pod number, pod weight, seed weight, seed size, shelling percentage and harvest index. Most heritability estimates for N2 fixation and related traits were moderate and high ranging from 0.33 to 1.00 except one low estimate (0.33). Most heritability estimates for agronomic traits were also moderate and high ranging from 0.37 to 1.00 except two low estimates (0.37 and 0.50). High heritability estimates in the F5 generation indicated that genetic variations of this population after divergence selection in the F4 generation are conserved and further selection for N2 fixation and agronomic traits in this population is possible when individual characters are considered. High and positive correlations among fixed nitrogen and its components were found indicating that increase in one trait can increase others. Correlations between N2 fixation and pod yield were low and varied depending on crosses suggesting that selection for both N2 fixation and pod yield is essential.
PDF Abstract XML References Citation
How to cite this article
J. Sikinarum, P. Jaisil, S. Jogloy, B. Toomsan, T. Kesmala and A. Patanothai, 2007. Heritability and Correlation for Nitrogen (N2) Fixation and Related Traits in Peanut (Arachis hypogaea L.). Pakistan Journal of Biological Sciences, 10: 1956-1962.
DOI: 10.3923/pjbs.2007.1956.1962
URL: https://scialert.net/abstract/?doi=pjbs.2007.1956.1962
DOI: 10.3923/pjbs.2007.1956.1962
URL: https://scialert.net/abstract/?doi=pjbs.2007.1956.1962
INTRODUCTION
The residual benefits of symbiotic N2 fixation by leguminous species in association with soil rhizobium to preserve soil fertility and crop productivity are well recognized. However, the utilization of leguminous species as green manure is limited for many reasons. The cultivation of non-grain legumes such as Sesbania spp. or others during follow period requires long duration of crop growth (60 days or longer) that is not suitable for many cropping systems and also needs additional phosphorus fertilizer as the starter and labor for incorporation of the stover into the soils. Farmers do not adopt green manure under rain-fed upland conditions. Furthermore, these species do not provide immediate cash for poor farmers and they are reluctant to adopt these leguminous species to improve their soils (McDonagh et al.,1995; Toomsan et al., 1995).
Grain legumes are more advantageous than non-grain legumes in crop rotation in that they can supply both food and soil improvement. However, the residual benefits of grain legumes are dependent upon the amount of fixed nitrogen left in stover after removal of marketable pods. Positive nitrogen balance is important for replenishing nitrogen into the soil when economic yield is harvested (van Kessel and Hartley, 2000). Peanut is the most promising grain legume for this purpose because after pods are removed it provides sufficient amount of nitrogen in stover that is beneficial to succeeding crops such as lowland rain-fed rice, upland rain-fed rice, field maize, sugarcane and cassava. This can provide sustainable means for crop production. However, some newly-released cultivars with high nitrogen harvest index could give negative balance of soil nitrogen (Bell and Wright, 1994). As N2 fixation competes with developing pods for assimilates, it is difficult for breeders to combine high N2 fixation with high pod yield and the only advantage of high N2 fixation has been an increase in the nitrogen in the residues (Redden, 2000).
The effective selection for traits under improvement depends on sufficient additive genetic variation of the traits that are expressed as heritability. Few studies so far have been conducted on the inheritance of N2 fixation and related traits in peanut. Arrendell et al. (1985) found that broad sense heritability estimates for nodule number, nodule weight, nitrogenase activity, shoot weight and fruit weight were moderate to high. Nigam et al. (1985) also reported predominant nature of non-additive genetic variances for nitrogenase activity and other traits. Provorov and Tikhonovich (2003) in their review reported high heritability of nitrogenase activity in many leguminous crops, suggesting that selection for this activity may be highly effective. For the inheritances of agronomic characters, fairly high heritability estimates for seed number, pod length and maturity index were observed, but heritability estimates for yield per plant was relatively low (Ali and Wynne, 1994; Ali et al., 1995).
Genetic relationships among traits are also important when simultaneous selection of multiple traits is to be carried out. High N2 fixing varieties should provide higher pod yield as well as high quality of stover. Close associations among nitrogenase activity and related traits such as nodule number, nodule mass, total nitrogen, shoot weight and root weigh were observed (Nigam et al., 1985; Arrendell et al., 1985). Arunachalam et al. (1984) reported that nitrogenase activity and nodule mass were the most important characters determining the relative performance of cultivars for high nitrogen fixation. Recently, Bado et al. (2006) observed significant correlation (p<0.05, R2 = 0.94) between total N yield of peanut and N derived from the atmosphere.
As breeding for high N2 fixation is the main objective of the ongoing peanut breeding program at Khon Kaen University, a number of F4 progenies from several crosses have been subjected to divergence selection for both high and low performances of direct N2 fixation and its surrogate traits (Pimratch et al., 2004b). The selected progenies were evaluated for heritability of N2 fixation and related traits in the F5 generation. We hypothesized that genetic variations for N2 fixation and related traits in this peanut population after selection are conserved in the F5 generation. The objectives of this study were to estimate broad sense heritability for N2 fixation and its related traits in peanut and evaluate correlation among the traits.
MATERIALS AND METHODS
Plant materials: Three peanut lines (PI 268770, PI 269109 and PI 152133) selected for high top dry weight, number of nodules and nodule weight from previous evaluation (Toomsan et al., 1991) and two high yielding cultivars (KKU 1 and KKU 72-1) in Thailand were crossed in 5x5 diallel design without reciprocals. The resulting F1 hybrids were grown for seed increase. The F2 crosses were further evaluated for combining ability and heritability for the characters related to N2 fixation and agronomic traits and the results were reported elsewhere (Phudenpa et al., 2004).
Six crosses (KKU 1xPI 152133, PI 268770xPI 269109, KKU 1xKKU 72-1, PI 268770xKKU 72-1, KKU 1xPI 269109 and KKU 72-1x PI 269109) with high general performance were selected and the randomly chosen 20 F2 families of each cross were planted for seed increase in F3 generation. The F4 seeds were evaluated for heritability within crosses and the results were reported previously (Pimratch et al., 2004b).
Based on the evaluation in the F4 generation, the F4 families were subjected to divergence selection for both high and low performance (bi-directional selection). The numbers of selected families for all crosses were not exactly equal, ranging from 10 to 14 families with the total number of 74 families and then they were available for evaluation in the F5 generation.
Experimental procedures: The experiment was conducted at the agronomy farm of Khon Kaen University in the rainy season 2003 with supplemental irrigation under limited soil nitrogen conditions. Lime (CaO) at the rate of 625 kg ha-1 was incorporated into the soil during soil preparation. The 74 families were laid out in a Randomized Complete Block Design (RCBD) with four replications. A non-nodulating line (referred to as non-nod) was also included as a reference plant for nitrogen determination. The plot size was three-row plot with 2 m in length and spacing of 50 cm between rows and 20 cm between plants within a row. Seeds were treated with captan at the rate of 10 g kg -1 seed to control seedling blight disease. The seeds were over-planted and later thinned to one plant per hill at seven days after emergence. Chemical fertilizers as P2O5 and K2O at the rates of 56.25 and 37.5 kg ha-1, respectively, were applied at 20 days after planting and gypsum (CaSO4) at the rate of 312.5 kg ha-1 was applied at 45 days after planting. Neither nitrogen fertilizer nor rhizobium inoculation was applied to the crop because the experimental site had history of peanut cultivation in previous years and peanut did not respond to the inoculation (Toomsan et al., 1988). Weeds, insects and diseases were adequately controlled during crop growth.
Ten plants in the middle row of each plot were harvested at maturity. Care had been taken to remove nodules from the soil as many as possible. Marketable mature pods were removed and air-dried. Plants were cut at crown level, roots after removal of the nodules were discarded and above ground parts (stems and leaves)and nodules were oven-dried at 80° C for 48 h. Dried pods were counted, shelled and then data were recorded for top dry weight, nodule dry weight, pod number, seed number and 100 seed weight on per plant basis. The derived data were also calculated for harvest index and shelling percentage.
To determine fixed nitrogen, dry aerial parts (stems and leaves) were chopped into small pieces and mixed thoroughly for individual samples. Small amounts of samples were taken from the bulked samples of 10 plants and ground for nitrogen determination by micro-Kjeldahl method (Black, 1965). Fixed nitrogen was calculated as;
Fixed nitrogen = total nitrogen of each genotype-nitrogen in reference plant (non-nod)
Data analysis: The data were subjected to analysis of variance using random statistical model as follows;
Yij = μ + ri + Fj + E ij,
where Yij, μ, ri, Fj and Eij represent observed value, overall mean, replication effect, family effect and error associated with observed value, respectively.
Duncan’s multiple range test was used to compare means (Gomez and Gomez, 1984).
Heritability in broad sense (H) was computed as suggested by Fehr (1987).
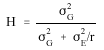 |
where Ó2G and Ó2E represent the variance components due to family and experimental error, respectively.
Heritability estimates were grouped as high (>0.50), moderate (0.20-0.50) and low (<0.20) as suggested by Stansfield (1986). Simple correlation coefficients were calculated using observed values of the plots. Most statistical analyses were performed using MSTAT-C package (Bricker, 1989), except for the partitioning of variance components and the calculation of heritability estimates which were performed manually using Microsoft Excel program.
RESULTS AND DISCUSSION
Heritability: Genotypes differences were highly significant (p<0.01) for all N2 fixation and related traits and all agronomic traits, confirming the presence of variability in the genetic materials (data not reported). Previous studies also revealed significant differences in their parents for these traits (Pimratch et al., 2004a). The significant differences among progenies allowed us to further process the data to determine the heritability. Estimates of heritability for N2 fixation and related traits ranged from 0.33 to 1.00 (Table 1). Most heritability estimates were high (>0.50) except one moderate heritability estimate (0.33) for leaf color score in the cross PI 268770xKKU 72-1. As the progenies evaluated were previously subjected to selection for high and low performance for nitrogen fixation and pod yield in stead of random selection, the more precise term used should be repeatability. Heritability is referred to either random segregating or naturally available population (Fehr, 1987).
Similar results of relatively high broad-sense heritability estimates have been reported for nitrogenase activity (0.60 to 0.82) and shoot dry weight (0.63 to 0.85) (Arrendell et al., 1986). However, Arrendell et al. (1988) reported very low realized heritability estimates for fresh plant weight (0.01), nodule number (0.05) and dry weight (0.06) and acetylene reduction (0.31) because of the insufficient control of environmental variation. The contrasting results might be due to differences in experimental conditions and materials used. Denison et al. (1985) found in soybean that variations in N2 fixation and related traits were affected by stresses and availability of soil nitrogen. In high available soil nitrogen plants were less dependent on fixed nitrogen and available soil nitrogen can alter relationships between nitrogen fixation and related traits. Present results would support the conclusion that evaluation of nitrogen fixation requires low soil nitrogen conditions and well-managed experiment.
Predominance of high heritability estimates for all traits in all crosses indicated that genetic variability is conserved in the F5 generation and divergence selection in the F4 generation had no significant effect on heritability estimates. These traits were highly heritable and less affected by environmental conditions under insufficient soil nitrate. Selection for these traits would be more effective and successful in advanced generations or even in early generation without going for further progeny testing.
| Table 1: | Heritability estimates for Leaf Color Score (LCS), Nodule Dry Weight (NDW), Shoot Dry Weight (SDW), Total Nitrogen (TN) and Fixed Nitrogen (FN) of six peanut crosses in the F5 |
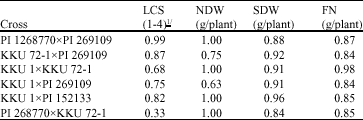 | |
| 1/ = Pale yellow ; 4 = Dark green | |
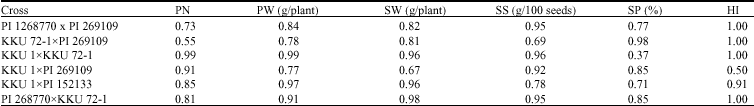
| Table 2: | Heritability estimates for Pod No. (PN), Pod Weight (PW), Seed No. (SN), Seed Weight (SW), Seed Size (SS), Shelling Percentage (SP) and Harvest Index (HI) of six peanut crosses in the F5 generation |
 | |
| Table 3: | Correlation coefficients between pairs of characters related to nitrogen fixation of six peanut crosses in the F5 generation |
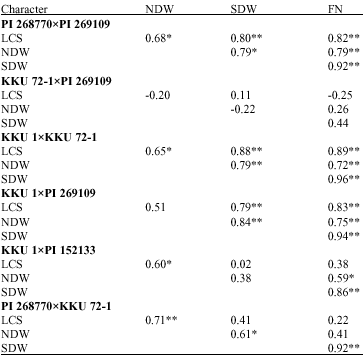 | |
*,** significant at 0.05 and 0.01 probability levels, respectively; Leaf Color Score (LCS), Nodule Dry Weight (NDW), Shoot Dry Weight (SDW) and Fixed Nitrogen (FN) | |
Similar to those for N2 fixation and related traits, the heritability estimates for all agronomic characters were moderate to high ranging from 0.37 to 1.00 (Table 2). Most of them were high (>0.50) except for shelling percentage in the cross KKU 1xKKU 72-1 (0.37). Earlier studies pertaining to heritability for agronomic traits in peanut have indicated variable results. Relatively high heritability estimates for both broad sense and narrow-sense for and pod length but relatively low for pod yield per plant were previously reported (Ali and Wynne, 1994; Ali et al., 1995). Green et al. (1983) reported moderate heritability estimates for yield and fruit traits, suggesting selection in later generations. The relatively high heritability estimates in this study should be resulted from genetic divergence in parental materials and the conservation of genetic variability after divergence selection in the F4 generation.
Correlation between traits: Significantly moderate to high correlation coefficients were found for all correlation combinations in the crosses PI 268770xPI 269109 (r = 0.68* to 0.92**) and KKU 1xKKU 72-1 (r = 0.65* to 0.94**) (Table 3). Significant correlation coefficient was not found in the cross KKU 72-1xPI 269109. Most correlation coefficients were highly significant and positive in the cross KKU 1xPI 269109 except for the correlation between nodule dry weight and leaf color score. Correlation coefficient between leaf color score and nodule dry weight was significant in the cross KKU 1xPI 152133. Correlation coefficients between fixed nitrogen and nodule dry weight and between fixed nitrogen and top dry weight were also significant and positive. In the cross PI 268770xKKU 72-1, Leaf color score was positively correlated with nodule dry weight. Nodule dry weight was also positively correlated with top dry weight and top dry weight and fixed nitrogen were well associated positively. In general, fixed nitrogen had strong, positive and consistent relationship with top dry weight across the crosses except in the cross KKU 72-1xPI 269109.
All the traits were generally well associated, it is interesting to note here that degrees of associations among N2 fixation and its components varied depending on crosses. This might be due to the differences in genetic backgrounds of their parents. Stalker et al. (1994) found that total dry matter accumulation was independent of root nodule or rate of fixation. Thus, correlations for these characters were not consistent across the crosses. However, the correlations between N2 fixation and its surrogate traits especially shoot dry weight were high in most crosses. This might be due to the close association between leaf nitrogen and carbon assimilation (Sinclair et al., 1993). Shoot dry weight is a good indicator for N2 fixation and should be use as a selection criterion. Because larger biomass crops require more nitrogen, N2 fixation will be increased with as biomass yield is increased (Herridge and Rose, 2000). However, this approach might be not necessary to increase pod yield, but truly improves capacity of N2 fixation sufficient for nitrogen requirement of larger crops.
Breeding for improved N2 fixation has not proven the end unto itself and improved yield and agronomic traits should be considered. Associations between traits related to nitrogen fixation and traits related to pod yield were studied to determine the consequence of selection.
| Table 4: | Correlation coefficients between pairs of agronomic characters and characters related to nitrogen fixation of six peanut crosses in the F5 generation |
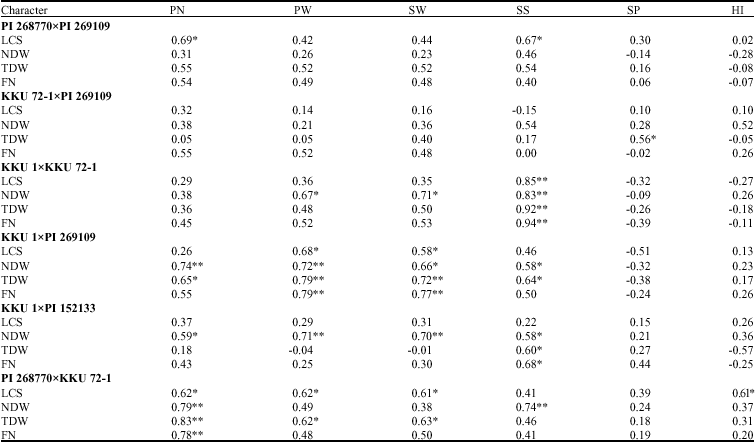 | |
*,** significant at 0.05 and 0.01 probability levels, respectively; Pod No. (PN), Pod Weight (PW), Seed Weight (SW) and Seed Size (SS), Shelling Percentage (SP), Harvest Index (HI), Leaf Color Score (LCS), Nodule Dry Weight (NDW), Total Dry Weight (TDW) and Fixed Nitrogen (FN) | |
Correlations between N2 fixations parameters and agronomic characters varied, depending on crosses (Table 4). In the cross PI 268770xPI 269109, most correlation coefficients were not significant and significant and positive correlation coefficients were found between leaf color score and pod number (r = 0.69*) and between leaf color score and seed size (r = 0.67*). Similar results were observed in the cross KKU 72-1xPI 269109 in which only correlation coefficient between top dry weight and shelling percentage (r = 0.56*) was significant and positive. High and positive associations were found between seed size and all nitrogen fixation parameters (r = 0.83** to 0.94**) in the cross KKU 1xKKU 72-1 and nodule dry weight was also correlated positively with pod weight (r = 0.67*) and seed weight (r = 0.71*). In the cross KKU 1xPI 269109, pod weight and seed weight were both positively correlated with all traits related to nitrogen fixation. Nodule dry weight and top dry weight were also positively correlated with pod number and seed size. In the cross KKU 1xPI 152133, seed size was positively correlated with nodule dry weight, top dry weight and fixed nitrogen and nodule dry weight was also positively correlated with pod number, pod weight and seed weight. In the cross PI 268770xKKU 72-1 pod number was positively correlated with all nitrogen fixation parameters. Pod weight and seed weight were positively correlated with leaf color score and top dry weight. Seed size was positively correlated with nodule dry weight and harvest index was positively correlated with leaf color score.
Implications for breeding for N2 fixation: The associations between traits related to N2 fixation and traits related to pod yield were not as high as the associations among each group of characters and the associations were low for shelling percentage and harvest index. This might be due to the fact that higher portion of fixed nitrogen was partitioned to shoot rather than to seed. Residual benefits of the stover after the removal of marketable pods on soil N fertility should result. However, N2 fixation may be low in high nitrate soils and should be future priority of breeding programs. Selection for nitrate tolerance should increase percent nitrogen under non-limited soil nitrogen conditions because it may be impossible to produce a legume solely dependent upon N2 fixation for growth (Herridge and Rose, 2000). Burias and Planchon (1990) found in soybean that seed yields of F4 derived lines were significantly correlated with N2 fixation of their parents and also correlated with nodule volume and nodule weight particularly at each cross level, suggesting possibility to improve yield.
Present results indicate that it is possible and effective to improve N2 fixation in this peanut population. Shoot dry weight is the best choice of characters alternative to direct N2 fixation for screening of large samples under limited soil nitrogen conditions. Simultaneous improvement of N2 fixation and pod yield is, in lesser extent, also possible and selection for both N2 fixation and pod yield is essential because low association between traits and the associations varied depending on crosses. This information should help breeders to formulate appropriate breeding strategies for improving N2 fixation and pod yield.
REFERENCES
- Ali, N. and J.C. Wynne, 1994. Heritability estimates and correlation studies of early maturity and other agronomic traits in two crosses of peanut (Arachis hypogaea L.). Pak. J. Bot., 26: 75-82.
Direct Link - Arrenddell, S., J.C. Wynne, G.H. Elkan and T.G. Isleib, 1985. Variation for nitrogen fixation among progenies of a virginia ✕ spanish peanut cross. Crop Sci., 25: 865-869.
CrossRefDirect Link - Bell, M.J. and G.C., Wright, 1994. The N2-fixation capacity of peanut cultivars with differing assimilate partitioning characteristics. Aust. J. Agric. Res., 45: 1455-1468.
Direct Link - Bado, B.V., A. Bationo and M.P. Cescas, 2006. Assessment of cowpea and groundnut contributions to soil fertility and succeeding sorghum yields in the Guinean Savannah zone of Burkina Faso (West Africa). Biol. Fertil. Soils, 43: 171-176.
Direct Link - Burias, N. and C. Planchon, 1990. Increasing soybean productivity through selection for N-fixation. Agron. J., 82: 1031-1034.
Direct Link - Fehr, W.R., 1987. Principles of Cultivar Development. Vol. 1, Macmillan Publishing Co., New York.
Direct Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons Inc., Hoboken, New Jersey, ISBN: 978-0-471-87092-0, Pages: 704.
Direct Link - Herridge, D. and I., Rose, 2000. Breeding for enhanced nitrogen fixation in crop legumes. Field Crops Res., 65: 229-248.
Direct Link - McDonagh, J.F., B. Toomsan, V. Limpinuntana and K.E. Giller, 1995. Grain legumes and green manures as pre-rice crops in Northeast Thailand: 1 Legume N 2-fixation, production and residual nitrogen benefits to rice. Plant Soil, 177: 111-126.
Direct Link - Phudenpa, A., S. Jogloy, B. Toomsan, S. Wongkaew, T. Kesmala and A. Patanothai, 2004. Heritability and phenotypic correlation of traits related to N2-fixation and agronomic traits in peanut (Arachis hypogaea L.). Songklanakarin J. Sci. Technol., 26: 317-325.
Direct Link - Pimratch, S., S. Jogloy, B. Toomsan, P. Jaisil, J. Sikhinarum, T. Kesmala and P. Patanothai, 2004. Evaluation of seven peanut genotypes for nitrogen fixation and agronomic traits. Songklanakarin J. Sci. Technol., 26: 295-304.
Direct Link - Pimratch, S., S. Jogloy, B. Tomsan, P. Jaisil, T. Kesmala and A. Pathanothai, 2004. Heritability and correlation for nitrogen fixation and agronomic traits of peanut (Arachis hypogaea L.). Songklanakarin J. Sci. Technol., 26: 305-315.
Direct Link - Provorov, N.A. and I.A. Tikhonovich, 2003. Genetic resources for improving nitrogen fixation in legume-rhizobium symbiosis. Genet. Resour. Crop Evol., 50: 89-99.
Direct Link - Sinclair, T.R., J.M. Bennett and K.J. Boote, 1993. Leaf nitrogen content, photosynthesis and radiation use efficiency in peanut. Peanut Sci., 20: 40-43.
Direct Link - Toomsan, B., J.F. McDonagh, V. Limpinuntana and K.E. Giller, 1995. Nitrogen fixation by groundnut and soybean and residual nitrogen benefits to rice in farmers-fields in Northeast Thailand. Plant Soil, 175: 45-56.
Direct Link - Van Kessel, C. and C. Hartley, 2000. Agricultural management of grain legumes: Has it led to an increase in nitrogen fixation? Field Crop Res., 65: 165-181.
Direct Link