
M. H. Mirmomeni
Department of Biology, Faculty of Science, Razi University, Kermanshah, Iran
A. Hosseinzadeh Colagar
Department of Biology, Faculty of Basic Science, Mazandaran University, Babolsar, Iran
S. Ghazaey
Department of Biology, Faculty of Science, Razi University, Kermanshah, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 10 | Page No.: 1562-1570







ABSTRACT
In this study two control isolates of Salmonella enteritidis, RTCC1623 and RTCC1624, were obtained from the institute of Razi (Karaj-Iran) and 14 strains were isolated from poultry samples in Kermanshah province of Iran, according to a standard protocol. These isolates were confirmed by PCR amplification of SefA gene fragments. Results showed that, 6 isolates of 14 isolates of Salmonella which their biochemical tests were positive contain 511 bp amplified fragments of the SefA gene. In other purpose, to correlating the presence of plasmids with antibiotic resistance and protein pattern, plasmid DNA was isolated before and after plasmid curing by using the alkaline lysis method. Strains of S. enteritidis contain seven different plasmid profiles (P1-P7) which were characterized by antibiotic resistance and protein pattern. Our observed showed, there was a high molecular weight plasmid with Rf 0.17 in all strains and the frequency of other plasmids was low. The plasmid with Rf about 0.2 is responsible for resistance to Cephalothin and the isolates that lost it were susceptible to this antibiotic. All strains, 100%, were resistant to ampicillin before and after curing of strains. According to present findings, PCR is a rapid and sensitive method for typing of S. enteritidis and plasmid profiling; antibiotic resistance and protein pattern are suitable methods for subtyping of S. enteritidis isolates. No direct correlation was found between plasmid contents, antibiotic resistance patterns and protein profiles of local S. enteritidis isolates.
PDF Abstract XML References Citation
How to cite this article
M. H. Mirmomeni, A. Hosseinzadeh Colagar and S. Ghazaey, 2007. Molecular Study of Salmonella enteritidis in Poultry Samples by PCR, Plasmid Curing, Antibiotic Resistance and Protein Pattern Analysis. Pakistan Journal of Biological Sciences, 10: 1562-1570.
DOI: 10.3923/pjbs.2007.1562.1570
URL: https://scialert.net/abstract/?doi=pjbs.2007.1562.1570
DOI: 10.3923/pjbs.2007.1562.1570
URL: https://scialert.net/abstract/?doi=pjbs.2007.1562.1570
INTRODUCTION
Salmonellae are among the most important agents of food-borne infections. Poultry, poultry products, cattle and dairy products are the major sources of Salmonella contaminated food products that cause human salmonellosis (Wegener et al., 1997). Salmonellosis caused by Salmonella enterica serovar enteritidis (S. enteritidis) is a disease affecting birds, mammals and humans. Therefore it is considered as a zoonosis (Kid et al., 1996). It has been said that the worldwide increase in S. enteritidis outbreaks is due to increased virulence. Virulence varies among different strains (Poppe et al., 1993a) and a specific plasmid has been suggested (Christensen et al., 1992; Parry, 1989). It is postulated that S. enteritidis colonized ovaries and oviducts of chickens and subsequently contaminates eggs as them form (Humphrey et al., 1991; Gast and Beard, 1990; Shivaprasad et al., 1990). S. enteritidis contaminates the content of intact eggs and is the major egg-associated human pathogen (Baulmer et al., 2000; Keller et al., 1997; Mead et al., 1999).
The PCR represents a major advance in terms of the speed, sensitivity and specificity of diagnostic methods. It has been increasingly used to identify several bacterial species from food and is perquisite for control of S. enteritidis infection in poultry (Stone et al., 1994). Nonselective and/or selective enrichment combined with PCR have been applied to detection of many bacterial pathogens (Candrian, 1995; Schrank et al., 2001) to improve sensitivity and dilute PCR inhibitory substances (Fluit et al., 1993). The objective of the present study was to establish a specific, sensitive and rapid PCR protocol for the detection of S. enteritidis a good candidate for identification of S. enteritidis by PCR is SefA gene. This gene codes for a major subunit protein of a novel fimbrial structure on the surface of S. enteritidis, designated Sef14 and is considered to be diagnostic significance (Turcotte and Woodward, 1993).
Control of S. enteritidis infection in poultry required intensive studies of pathogen-markers needed in determining the relatedness among S. enteritidis strains recovered during different epidemiological investigations. The study of relatedness among S. enteritidis strains is a perquisite segment in developing vaccines against S. enteritidis infection. Phenotypic markers used in previous studies on S. enteritidis included antimicrobial susceptibility testing (Corpus et al., 2004) phage typing (Tsen and Lin, 2001) recently works on phenotypic markers of S. enteritidis have been supplemented by genetic characterization including plasmid profiling and protein pattern analysis (Icgen et al., 2002).
Plasmid profiling is one of the genetic methods which have been used to differentiate strains of S. enteritidis (Olsen et al., 1993). S. enteritidis frequently possesses plasmids of different sizes and roles. The most frequent and best characterized is the virulence plasmid with molecular mass around 55 Kb (Buisan et al., 1994; Helmuth et al., 1985). Besides the virulence plasmid, S.enteritidis can harbour plasmids of low molecular mass. Their biological roles are nearly unknown and only a few of them have been characterized in detail. Some of these plasmids most probably associated with antimicrobial sensitivity (Brown et al., 1994). S. enteritidis is more susceptible to available antimicrobial agents than other Salmonella serotypes (Lee et al., 1994). The characteristic migration patterns of proteins during SDS-PAGE have been used for subdividing Salmonella serotypes as such patterns yield information on how such organisms are related (Helmuth et al., 1985; Poppe et al., 1993b). To correlate the presence of plasmids to the other characteristics 6 strains that harbour virulence plasmid either on its own or in addition to other plasmids were chosen arbitrarily the present study investigates the effects of loss of specific plasmids on the resistance to antimicrobial agents and protein profiles in this isolates.
MATERIALS AND METHODS
Preparation of samples and grows condition: Forty S. enteritidis isolates were obtained from poultry samples for example chicken meat and liver and egg. Five grams of poultry homogenized with 25 mL of Selenite F broth (Enrichment broth) was incubated at 37°C for 24 h to allow for bacterial grows and then cultured in Salmonella Shegella/ S.S agar selective medium to select the colorless single colonies. All the strains were identified in laboratory according to a standard protocol (Ward et al., 1987) including biochemical standard tests. Fourteen isolates of S. enteritidis (E1-9, M1-3 and L1-2, the letters E, M and L referring to egg, meat and liver origins, respectively), were prepared for PCR. In this study the strains were maintained as frozen stocks in Leuria broth/LB supplemented with 20% glycerol at 70°C.
DNA extraction: Chromosomal DNA isolated by boiling method, ten colonies of bacteria from S.S agar medium was suspend in a micro centrifuge tube with a capacity of 1.5 mL with 300 μL of DNase-RNase-free distilled water by vortexing. Heating of micro centrifuge tube in boiling water for 15 min, suspension was centrifuged for 5 min at 6,000 g. The supernatant was extracted carefully and was ready for electrophoresis.
Plasmid DNA was isolated using the STET boiling method described by Sambrook et al. (2001).
PCR: The oligonucleotide primers used for the specific detection of the SefA gene, SefA-F (5’ GCGCCATGGGTAAATCAGCATCTGC 3’) and SefA-R (5’ CCCTCTAGAAGGTTTTGATACTGCT 3’) described by Kisiela et al., (2002). The SefA DNA fragment (511 bp) was amplified as follow: 25 cycles of denaturation (94°C, 3 min), annealing (52°C, 1 min), elongation (72°C, 1.5 min). Amplification products were visualized by agarose gel electrophoresis (Samobrook et al., 2001). The sensitivity of the PCR reaction was determined for S.enteritidis isolates using the non-Salmonella pool.
Antibiotical susceptibility testing: The antibiotical susceptibility assays were performed by standard disc diffusion method on Muller-Hinton agar, as described in the NCCLS guidelines. The following disc antibiotics and the respective concentration, purchased from Padtan Teb Co. of Iran, including: Ampicillin (10 μg); Kanamycin (10 μg); Cephalothin (30 μg); Ciprofloxacin (5 μg); Amikacin (30 μg); Cefotaxime (30 μg); Ceftizoxime (30 μg); Cefalexin (30 μg); Carbencillin (100 μg); Nitrofurantoin (300 μg); Norfloxacin (10 μg); Nalidixic acid (30 μg); Streptomycine (10 μg); Getnamicin (10 μg); Trimethoprim-sulfamethazole (75 μg).
Protein extraction: Preparation of total protein content was performed by employing alkaline lysis solution (contain 50 mM glucose, 10 mM EDTANa2 pH 8.0, 25 mM tris-HCl; pH 8.0) digestion. Protein content of isolates were analyzed with a dis continues SDS-PAGE system (Laemmli, 1970) which contain 12% separating gel and a 5% stacking gel. Gels were stained with Coomassie blue using the method described by Tsai and Frasch (1982).
Plasmid curing: An SDS solution (10% w/v, pH 7.4) was added to LB (double strength) and the final volume was made up to 10 mL in MacCartney bottles with distilled H2O to give 5, 4, 3, 2, 1 and 0.5% SDS in the LB (normal strength). An inoculum of 100 μL (1.2832 ’ 109 cells mL-1) of the wild-type was then used to seed the SDS containing LB and the cultures were incubated overnight with shaking at 45°C and sub cultured for 6 days (Mansi et al., 2000). After which a dilution form each culture was plated on to S.S agar. To select the derivatives that had lost antibiotic resistance the colonies were transformed by tooth picking on to Muller Hintone agar media.
RESULTS
PCR specificity and sensitivity: Chromosomal DNA were extracted from Salmonella strains which was determined in 14 poultry products by biochemical tests. The PCR reaction performed and single band of 511 bp amplified by SefA-F/ SefA-R primers (Fig. 1A). Confirmation of PCR product from positive isolates performed by AvaII restriction enzyme and two 217 and 281 bp fragments visible (Fig. 1B). Results shown that, 6 isolates of fourteen isolates of S. enteritidis which are positive biochemical tests, contain these fragments.
Plasmid profiles: The plasmid profile has been used in epidemiological studies of S. enteritidis. In 14 isolates of S. enteritidis seven different plasmid profiles, P1-P7, were identified (Table 1). The band numbers for these plasmid profiles were 1 to 7 with Rf value ranging from 0.05 to 0.66. For simple subdivision of S. enteritidis isolates we calculated the Rf value for each band of plasmid profiles. We found one high molecular weight plasmid with Rf 0.17 that was present in all isolates. Isolates harbour multiple types of low molecular weight plasmids.
Plasmid curing: The isolates of E1, E3, E8, M2 and L1-2 were used in the curing experiments. Optimization the assay was done by various concentration of SDS. We found that reveal concentration for curing multiple types of plasmid bands is 2% SDS. In this condition we succeed to cure a lot of plasmid bands of isolates (Fig. 2).
Antibiotic susceptibility: Our observation showed that 100% of isolates were resistant to Ampiciline. Eighty five percent of S. enteritidis isolates were resistant to Streptomycin, 67% to Cephalothin and Cefotaxime and 17% of them to Ceftizoxime, Ciprofloxacin, Ciprofloxacin, Nalidixic acid and Norfloxacin (Table 2).
All the strains were susceptible to Carbenciline, Cephalexin. After curing the antibiotic sensitive derivatives of the isolates were tested. Changes in the antibiotic susceptibility patterns of the strains upon curing are presented in Table 2. Resistance to antimicrobial agents is changed by curing analysis and cured isolates were susceptible to a lot of antibiotics that have been resistant before curing experiment but after plasmid curing resistance to ampiciline was not changed. Antibiotic susceptibility tests proved valuable in differentiating strains of Salmonella spp. However, we did not find any correlation between S. enteritidis strains regarding their antibiotic susceptibility, which suggests that the tests cannot be used to distinguish among the strains.
Brown et al. (1994) detected a very low percentage (3.7 and 1%) of antibiotic resistance among the S. enteritidis strains they tested. the test of drug resistance, about all of the S. enteritidis strains showed resistance to various antibiotics examined; however, all strains that harbored only 36 MDa plasmid were sensitive to drug.
Protein pattern: Protein patterns of the 6 isolates before and after curing were demonstrated by SDS-PAGE (Fig. 3).
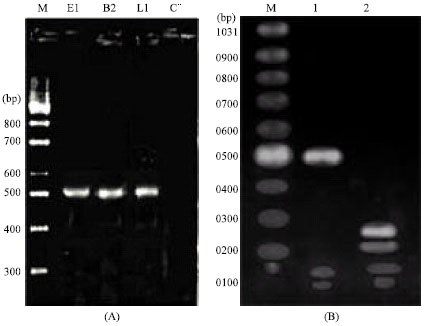 | |
| Fig. 1: | PCR product and confirmation: The SefA DNA fragment (511 bp) was amplified by SefA-F and SefA-R primers (A), Confirmation of PCR product from positive isolates performed by AvaII restriction enzyme and two 217 and 281 bp fragments visible (B). E1, M2 and L1 referring to egg, meat and liver origin of isolates, respectively. C– = negative control, M = DNA marker |
| Table 1: | Plasmid profiles of S. enteritidis isolates prepared by PCR |
 | |
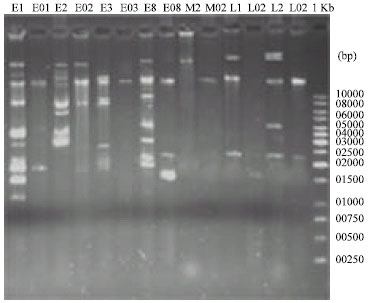 | |
| Fig. 2: | Plasmid profiling in non cured and cured S. enteritidis isolates: Plasmid curing performed by 2% SDS, Cured isolates names labeled by zero before number. 1 Kb is linear DNA marker |
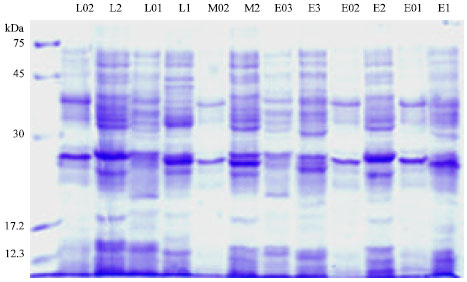 | |
| Fig. 3: | Total protein pattern in non cured and cured S. enteritidis isolates: SDS-PAGE performed in 10% resolving and 4% stacking gels, cured isolates names labeled by zero before number |
| Table 2: | Effect of plasmid curing on antibiotic susceptibility and protein pattern |
 | |
*AMP: Ampicillin, AMK: Amikacine, CEF: Cephalothin, CIP: Ciprofloxacin, CET: Ceftizoxime, CTX: Cefotaxime, GEM: Gentamycin, NIF: Nitrofurantoin, KAN: Kanamycin, NDA: Nalidixic acid, NOR: Norfloxacin, STR: Streptomycin, STX: Trimethoprim-sulfamethoxazole | |
The protein profile of S. enteritidis strains have been found to vary occasionally in conjunction with possession of plasmids. It was reported that a same profile was characteristic of six strains harbouring different plasmids (Helmuth et al., 1985). This analysis demonstrated that a lot of protein bands were same in six isolates and most of them were deleted by curing. For simple comparison of protein profiles of isolates we calculated Rf value of each band.
DISCUSSION
Biochemical typing of S. enteritidis isolates can be considered a first step in classification (Millemann et al., 1995). The application of PCR techniques had a revolutionary impact on the diagnosis of infectious disease. Because these techniques have the ability to detect or allow analysis of minute amount of microbial DNA sequences, they have emerged as a highly sensitive and specific method for identifying pathogens.
For further identification of S. enteritidis isolates obtained form PCR, Plasmid profiling is used as a traditional method for sub typing the isolates.
Plasmid profiling has been used frequently in typing of S. enteritidis despite the fact that most S. enteritidis strains contain only the serovar specific virulence plasmid (Threlfall et al., 1989; Pohl et al., 1991; Dorn et al., 1993; Rychlik et al., 1993; Brown et al., 1994). In the present study we isolated a high molecular weight plasmid with Rf 0.17 from all strains and the frequency of other plasmids was low. It was observed that the latter plasmids were detected mostly in multiresistant strains. Within strains about 35.7% of isolates contain only the high molecular weight plasmid with Rf 0.17 and cannot be further distinguished (Threlfall et al., 1989).
Plasmid profiling is therefore of rather questionable value in these cases. Low molecular weight plasmids were described even earlier (Threlfall et al., 1989). P1 profile was the major expression among this isolates. As there are strains of different plasmid profiles in different countries it is probable that there are different local sources of S. enteritidis infection within the continent. Several laboratories have reported the presence of a 54-57 kb virulence plasmid in most of the S. enteritidis strains studied (Helmuth et al., 1985; Bichler et al., 1994). We thought that the plasmids present in all of our strains (Rf 0.17) might be such 54-57 kb plasmid that Tsen and Lin in 2001 reported it with same 0.174 Rf value. It should be noted that most published data reporting plasmid size have been obtained from fragment size standards and interpreted by semi logarithmic interpolation. Considerable size errors can be generated, complicating comparisons of DNA fragments reported from different laboratories (Towner and Cockayne 1993). Thus, in this study, we used the migration distance or Rf value of each band to show the plasmid profile.
It is generally accepted that the metabolic burden imposed on the cell as a consequence of plasmid replication and gene expression diminishes the growth rate and, as such, plasmid-less cells are at an advantage with respect to fitness, as they grow faster (Bhattacharya and Dudey, 1995). In the absence of selective pressure, however, plasmids may survive by integrating into the genome, which is in the consequence of plasmid loss or integration into the host genome was apparent as it was accompanied by a slight but obvious change in the color and morphology of the colonies, (Holmes and Aucken, 1998). During the Course of growth, plasmid loss may be triggered in response to changes in environmental parameters such as temperature and pH (Gupta et al., 1995), changes in plasmid topology such as multimerization (Summers et al., 1991; Summers et al., 1993) or concatameric replication (Viret et al., 1991). Plasmid integration, on the other hand, is presumably triggered and catalyzed in a manner similar to that reported for site-specific recombination (Sherratt et al., 1995).
One important feature that is shared between genomic (chromosome) and plasmid DNA is their direct attachment to cell membrane (Brown, 1999) SDS, an anionic detergent that is widely used for the disruption of the cell membrane, may, at the right concentration, be capable of dislodging the indigenous plasmid free of its site of attachment which in turn leads to imperfect replication and unsuccessful segregation of the plasmid. Unlike strains that sustained damage to their chromosome as a result of the SDS treatment, cured strains, i.e., plasmid-less cells, can survive in a rich medium. The ability of SDS to dislodge plasmid molecules from their site may also account for the earlier observation that SDS prevents the conjugal transfer of R-plasmids (Tomas et al., 1986) as replication and subsequent segregation of these plasmids would be impaired owing to the lack of attachment sites. It is also possible for SDS, once it reaches the cytoplasm, to interfere with cellular metabolism through its ability to dissociate proteins into their respective subunits thus rendering some enzymes partially or fully inactive. Consequently, a plasmid can be lost if any of the inactivated enzymes is functionally associated with replication and/or segregation of the plasmid. In conclusion, the role portrayed for SDS as a curing agent in this study is largely due its ability to dislodge the plasmid from its site of attachment in the cell membrane and to a lesser extent, its capacity to denature proteins.
In the present study, our finding indicate that most strains were resistant to a wide rang of antimicrobial agents. Considering that 100% resistance to ampicillin was found in S. enteritidis strains containing 1 to 6 plasmids before and after curing, it was not possible to correlate the presence of these plasmids with the antibiotic resistance detected in some strains, but the isolates were susceptible to wide rang of antibiotics by curing with SDS. We thought that a plasmid with Rf about 0.2 is responsible for resistance to Cephalothin and the isolates that loss one was susceptible to this antibiotic. Antibiotic resistance in Salmonella is on the increase (Tassios et al., 1997). Although generally the resistance in Salmonella is not considered to be dependent on plasmids, resistance to antimicrobial agents is considered to be relatively unstable, because the majority of bacterial resistance factors are encoded by plasmids that are often transferable between strains and which may be dependent on selection pressure to be maintained (Olsen et al., 1993). Some examples of plasmid encoded resistances have been described (Vatopoulos et al., 1994; Gazouli et al., 1998). The function of low molecular weight plasmids in S. enteritidis is usually unknown although some indications on their contribution to phage typing results have been described (Gado et al., 1998; Fernandes et al., 2003). However, plasmids of higher molecular weight have been shown to affect the phenotype of S. enteritidis considerably, including their virulence (Helmuth et al., 1985; Buisan et al., 1994), phage type and/or antibiotic resistance (Vatopoulos et al., 1994) which could be a reason for the strains resistance to ampicillin. We did not find any other significant link between particular plasmids and resistance to antibiotics.
It has also been observed that antibiotic sensitivity differs for each serotype; even though, a general increase in number of resistant isolate has been observe this type of studies should contribute to the control of these infections as well as to raise the level of awareness about increased resistance to some antibiotics this should encourage the implementation of public health measures and the control of antibiotic use, mainly in animals (Tan et al., 1999).
Protein profiles of the isolates were changed by plasmid curing with SDS, that this result confirms accuracy of curing analysis. In conclusion, no direct correlation was found between antibiotic resistance patterns, plasmid contents and protein profiles of local S. enteritidis isolates. The heterogeneity of the plasmid profiles among our isolates is thought to reflect the sporadic and non-repetitive nature of the isolates.
REFERENCES
- Baulmer, A.J., B.M. Hargis and R.M. Tsolis, 2000. Tracing the origins of Salmonella outbreaks. Science, 287: 50-52.
Direct Link - Bhattacharya, S.K. and A.K. Dudey, 1995. Metabolic burden as reflected by maintenance coefficient of recombinant Escherichia coli over expressing target gene. Biotechnol. Lett., 17: 1155-1160.
Direct Link - Bichler, L.A., K.V. Nagaraja and B.S. Pomeroy, 1994. Plasmid diversity in Salmonella enteritidis of animal, poultry and human origin. J. Food Prot., 57: 4-11.
Direct Link - Brown, D.J., D.L. Baggesen, H.C. Hansen and M. Bisgaard, 1994. The characterization of Danish isolates of Salmonella enterica serovar enteritidis by phage typing and plasmid profiling. Acta Pathologica Microbiologica Immunologica, 102: 208-214.
Direct Link - Buisan, M., J.M. Rodriguez-Pena and R. Rotger, 1994. Restriction map of the Salmonella enteritidis virulence plasmid and its homology with the plasmid of Salmonella typhimurium. Microb. Pathog., 16: 165-169.
Direct Link - Candrian, U., 1995. Polymerase chain reaction in food microbiology. J. Microbiol. Metheds, 23: 89-103.
Direct Link - Christensen, P., E. Olsen, C. Hansen and M. Bisgaard, 1992. Characterization of Salmonella enterica serovar gallinarum and pullorum by plasmid profiling and biochemical analysis. Avian Histol., 21: 461-470.
Direct Link - Dorn, C.R., R. Silapanuntakul, E.J. Angrick and L.D. Shipman, 1993. Plasmid analysis of Salmonella enteritidis isolated from human gastroenteritis cases and from epidemiologically associated poultry flocks. Epidemiol. Infect., 111: 239-243.
Direct Link - Fernandes, S.A., C.R.T. Ghilardi, M.O. Tavechio Machado and C.C. Pignatari, 2003. Phenotypic and molecular characterization of Salmonella enteritidis strains isolated in Sao Paulo, Brazil. Rev. Inst. Med. Irob. S. Paulo., 45: 59-63.
Direct Link - Gado, I., V.G. Laszlo, B. Nagy, H. Milch, I. Drin, M. Awad-Masalmeh and J. Horvath, 1998. Phage restriction and the presence of small plasmids in Salmonella enteritidis. Zbl. Bakt. Int. J. Med. Microbiol., 287: 509-519.
Direct Link - Gast, R.K. and C.W. Beard, 1990. Production of Salmonella enteritidis-contaminated eggs by experimentally infected hens. Avian Dis., 34: 438-446.
CrossRefDirect Link - Gazouli, M., E. Tzelepi, A. Narkogiannakis, N.J. Legakis and L.S. Tzouvelekis, 1998. Two novel plasmids mediated cefotaxime-hydrolyzing β-lactamases (CTX-M-5 and CTX-M-6) from Salmonella typhimurium. FEMS Micribiol. Lett., 165: 289-293.
Direct Link - Gupta, R., P. Sharma and V.V. Vyas, 1995. Effects of growth environment on the stability of a recombinant shuttle plasmid in Escherichia coli. J. Biotechnol., 41: 29-37.
Direct Link - Humphrey, T.J., A. Baskerville, H. Chart, B. Rowe and A. Whitehead, 1991. Salmonella enteritidis PT4 infection in specific pathogen free hens: Influence of infecting dose. Vet. Rec., 1: 482-485.
Direct Link - Icgen, B., G.C. Gurakan and G. �zcengiz, 2001. Effects of plasmid curing on antibiotic susceptibility, phage type, lipopolysaccharide and outer membrane protein profiles in local Salmonella isolates. Food. Microbiol., 18: 631-635.
Direct Link - Keller, L.H., D.M. Schifferli, C.E. Benson, S. Aslam and R.J. Eckroade, 1997. Invasion of chicken reproductive tissues and forming eggs is not unique to Salmonella enteritidis. Avian Dis., 41: 535-539.
Direct Link - Kid, H., D.H. Read, R.P. Chin, A.A. Bickford, R.L. Walker and A. Ardans, 1996. Salmonella enteritidis phage type 4 infection in a commercial layer flock in Southern California. Bacteriologic and epidemiologic finding. Avian Dis., 40: 665-671.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Mansi, M., K.J. anderson, C.A Inche, L.K. Knowles and D.J. Platt, 2000. Isolation and curing of the Klebsiella pneumoniae large indigenous plasmid using sodium dodecyl sulphate. Res. Microbiol., 151: 201-208.
Direct Link - Mead, P.S., L. Slutsker, V. Dietz, L.F. McCaig and J.S. Bresee et al., 1999. Food-related illness and death in the United States. Emerg. Infect. Dis., 5: 607-625.
Direct Link - Millemann, Y., M.C. Lesage, E.C. Dancla and J.P. Lafont, 1995. Value of plasmid profiling, ribotyping and detection of IS200 for tracing avian isolates of Salmonella typhimurium and S. enteritidis. J. Clin. Microbiol., 33: 173-179.
Direct Link - Olsen, J.E., D.J. Brown, M.N. Skov and J.P. Christensen, 1993. Bacterial typing methods suitable for epidemiological analysis. Application in investigations of Salmonellosis among livestock. Vet. Q., 15: 125-135.
Direct Link - Pohl, P., P. Lintermans, M. Marin and M. Couturier, 1991. Epidemiological study of Salmonella enteritidis strains of animal origin in Belgium. Epidemiol. Infect., 106: 11-16.
Direct Link - Rychlik, I., F. Sisak and P. Lany, 1993. Differentiation of Salmonella enteritidis and Salmonella typhimurium by plasmid profile analysis and restriction endonuclease analysis of chromosomal DNA. Vet. Med. (Praha), 38: 433-439.
Direct Link - Schrank, I.S., M.A.Z. Mores, J.L.A. Costa, A.P.G. Frazzon and R. Sonicini et al., 2001. Influence of enrichment media and application of a PCR based method to detect Salmonella in poultry industry products and clinical samples. Vet. Microbiol., 82: 45-53.
Direct Link - Sherratt, D.J., L.K. Arciszewska, G. Bladely, G. Colloms, S. Grant, K. Leslie and R. McCulloch, 1995. Site specific recombination and circular plasmid chromosome segregation. Phil. Trans. R. Soc. Lond. Ser. B. Biol. Sci., 347: 37-42.
Direct Link - Shivaprasad, H.L., J.F. Timoney, S. Morales, B. Lucio and R.C. Baker, 1990. Pathogenesis of Salmonella enteritidis infection in laying chickens. I. Studies on egg transmission, clinical signs, fecal shedding and serologic responses. Avian Dis., 34: 548-557.
CrossRefDirect Link - Stone, G.G., R.D. Oberst, M.P. Hays, S. McVey and M.M. Chengappa, 1994. Detection of Salmonella serovars from clinical samples by enrichment broth cultivation-PCR procedure. J. Clin. Microbiol., 32: 1742-1749.
Direct Link - Tassios, T., A. Markkogiannakis, C. Vatopoulos, E. Katsanikou, N. Velonakis, J. Kremanstinou and J. Legakis, 1997. Molecular epidemiology of antibiotic resistance of Salmonella enteritidis during a 7 year Period in Greece. J. Clin. Microbiol., 35: 1316-1321.
Direct Link - Tsen, H.Y. and J.S. Lin, 2001. Analysis of S. enteritidis strains isolated form food-poisoning cases in Taiwan by pulsed field gel electrophoresis, plasmid profile and phage typing. J. Applied Microbiol., 91: 72-79.
Direct Link - Turcotte, C. and M.J. Woodward, 1993. Cloning, DNA nucleotide sequence and distribution of the gene encoding the SEF14 fimbrial antigen of S. enteritidis. J. Gen. Microbiol., 39: 1477-1485.
Direct Link - Vatopoulos, A.C., E. Mainas, E. Balis, E.J. Threlfall and M. Kanelopoulou et al., 1994. Molecular epidemiology of ampicillin-resistant clinical isolates of S. enteritidis. J. Clin. Microbiol., 32: 1322-1325.
Direct Link - Viret, J.F., A. Bravo and J.C. Alonso, 1991. Recombination dependent concatameric plasmid replication. Microbiol. Rev., 55: 675-683.
Direct Link