
Yelda Emek
Department of Biology, Faculty of Science and Arts,University of Adnan Menderes, 09010 Aydin, Turkey
Beng Erda
Department of Biology, Faculty of Science and Arts,University of Adnan Menderes, 09010 Aydin, Turkey
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 8 | Page No.: 1190-1194







ABSTRACT
An in vitro micropropagation method by somatic embryogenesis was developed for Gladiolus anatolicus (Boiss.) Stapf using leaves of in vitro shoots obtained from lateral buds. Lateral buds removed from sterilized fresh corms were placed on Murashige and Skoog (MS) medium supplemented with various concentrations of N6-benzyladenine (BA) for shoot culture establishment. The highest number of shoot per lateral bud explant was on MS medium supplemented with 2 mg L-1 BA (11.00±0.38). To induce somatic embryogenesis, leaves of in vitro shoots obtained from lateral buds were used as explant. Calli were obtained from middle and basal region of leaf explant cultured on MS basal medium supplemented with different concentrations of α-naphthaleneacetic acid: N6-benzyladenine (NAA:BA) ratio and without growth regulators. The highest rate of callus formation was obtained from basal part of leaves cultured on MS medium containing 5 mg L-1 NAA in darkness (80±0.41%). Creamy-white and friable calli produced numerous somatic embryos on MS basal medium supplemented with 0.1 mg L-1 BA within 4 weeks in light (On avarage 30 structures per callus). Well-developed somatic embryos were germinated on MS medium supplemented with 0.1 mg L-1 BA and reduced sucrose concentration (20 g L-1). On this medium 40% of the somatic embryos develepod into plantlets. Cormlet formation was observed on MS basal medium (30 g L-1 sucrose) containing same concentration of BA.
PDF Abstract XML References Citation
How to cite this article
Yelda Emek and Beng Erda, 2007. Somatic Embryogenesis from Leaf Explants of Gladiolus anatolicus (Boiss.) Stapf. Pakistan Journal of Biological Sciences, 10: 1190-1194.
DOI: 10.3923/pjbs.2007.1190.1194
URL: https://scialert.net/abstract/?doi=pjbs.2007.1190.1194
DOI: 10.3923/pjbs.2007.1190.1194
URL: https://scialert.net/abstract/?doi=pjbs.2007.1190.1194
INTRODUCTION
Natural propagation rate of many geophytes, including Gladiolus, is low. Plant micropropagation a novel biotechnological method for rapid mass production of plants, has been found valuable in the propagation of geophytes (Hussey, 1975, 1986; Krikorian and Kann, 1986). Somatic embryogenesis is the fastest plant micropropagation method and somatic embryos can also be encapsulated in various gels to form synthetic seeds that can be easily stored and transported long distances (Ghosh and Sen, 1994). Although there are many reports on Gladiolus micropropagation (Ziv et al., 1970; Ginzburg and Ziv, 1973; Dantu and Bhojwani, 1987, 1992; Kim et al., 1988; Kumar et al., 1999), only a few number of studies presented dealing with its somatic embryogenesis (Stefaniak, 1994; Tometsune et al., 1994; Remotti, 1995).
Gladiolus anatolicus (Boiss.) Stapf is a monocot belonging to the Iridaceae family, an endemic plant of Southwest Anatolia. Moreover, because of its striking appearance this spring flowering cormous species has a great potential to use as an ornamental. G. anatolicus would be of great importance for breeding programs to produce high quality Gladioli for flower market.
The present study is the first report on induction of somatic embryogenesis from callus cultures of G. anatolicus. The development of this protocol has opened up the possibility for large-scale clonal propagation of G. anatolicus.
MATERIALS AND METHODS
Plant material: In this study, leaves of in vitro shoots obtained from lateral buds of fresh corms of Gladiolus anatolicus (Boiss.) Stapf were used as explant.
Media preparation: All the experiments were maintained on semi-solid basal medium supplemented with various concentration of NAA and BA. Basal medium contained Murashige & Skoog (Murashige and Skoog, 1962,) (MS) mineral salts, 100 mg L-1 myo-inositol, 2 mg L-1 glycine, 0.5 mg L-1 nicotinic acid, 0.5 mg L-1 pyridoxine HCl, 0.1 mg L-1 thiamine HCl, 30 g L-1 sucrose, 8 g L-1 agar-agar (Sigma). Plant growth regulators were added prior to autoclaving as optional additives according to the experimental objectives. The pH of the media was adjusted to 5.8 with 1 N NaOH or HCl prior to autoclaving for 15 min at 121°C.
Culture conditions: Light-grown cultures were maintained at 24±2°C with illumination provided by cool white florescent lamps at 40 μE m-2 s-1 with a 16 h light period. Dark cultures were incubated at the same temperature. Subculturing was periodically carried out 4 weeks intervals.
Establishment of shoot cultures: Corms were collected from natural habitat before flowering period and were washed in running top water for 24 h and outer scales were removed. Corms were surface-sterilized with 70% EtOH for 17 min and with 4.5% sodiumhypochlorite for 20 min, consecutively and washed three times with sterilized distilled water. Lateral buds with approximately 3 mm edge were removed from sterilized corms.
For shoot proliferation, each lateral bud was placed with a surface in contact with MS basal medium containing various concentration of BA (0.5, 1 and 2 mg L-1). The cultures were maintained at 24±2°C in light.
Induction of somatic embryogenesis: To induce somatic embryogenesis, leaves of in vitro shoots obtained from lateral buds were used as explant. Leaf explants were cut transversely in to 10 mm lenght and base, middle and top region of leaves cultured with their abaxial surfaces in contact with the MS basal medium supplemented with or without BA: NAA (0:0, 0:1, 5:0, 5:1, 10:0, 10:1). The cultures were maintained at 24±2°C in the darkness. Creamy-white friable calli obtained from base and middle region of leaves were transferred to semi-solid MS basal medium supplemented with BA (0, 0.1, 0.5 or 1 mg L-1) for embryo differentiation and maturation. The terms of differentiation and maturation in this study denote the development of globular (early) somatic embryos into scutellar and coleoptilar stage. The cultures were maintained at 24±2°C in light.
After 8 weeks, well-developed somatic embryos were transferred to MS basal medium supplemented with or without 0.1 mg L-1 BA and the same media containing reduced sucrose concentration (20 g L-1) for embryo germination.
Plantlets were transferred to MS basal medium with or without 0.1 mg L-1 BA for cormlet formation.
Statistical analysis: All experiments were conducted with three replications consisting of four explants per 100 mL flask and were repeated twice. Data represented as mean±SE (standard error) or mean±SD (standard deviation).
RESULTS AND DISCUSSION
Sterile cultures were obtained in high proportion (95%) after two weeks of incubation that showed the successful sterilization procedure. Lateral buds cultured on MS medium supplemented with 1 mg L-1 BA underwent moderate swelling (after 3-4 days from culture initiation) at the base, followed by elongation of a single shoot after approximately 20-25 days of incubation. It is accordant with a report of Sen and Sen (1995) for Gladiolus cultivars. However, the single shoots were shown chlorosis and died off a short time later. Basal portion of the lateral buds cultured on MS basal media supplemented with 0.5 and 2 mg L-1 BA, became heavily swollen, followed by the induction of profuse shoot bud initials. Number of shoots of lateral buds inoculated on MS basal medium containing 2 mg L-1 BA was higher than MS basal medium supplemented with 0.5 mg L-1 BA, respectively and 11.00±0.38 and 7.96±0.41 (Fig. 1). The highest mean of maximum shoot length was also obtained in this medium. For this reason it can be stated out that the best medium is MS basal medium supplemented with 2 mg L-1 BA for shoot formation from lateral buds (Table 1).
After 6-8 weeks of culture initiation, leaves of obtained in vitro shoots were used for induction somatic embryogenesis. Callus formation was strongly affected by the region of leaf explant. Calli were initiated from base and middle region of leaves in darkness within 8 weeks. There was no callus formation from top region of leaves (Table 2). Probably a result of special growth pattern of a monocot leaf. The highest frequency of callus formation was obtained from base region of leaves in MS medium containing 5 mg L-1 NAA (80±0.41%). It was not observed any callus formation on medium without growth regulator. Moreover after the addition of BA into medium, callus formation rates were reduced. For this reason, it is accepted that 5.0 mg L-1 of NAA concentration is optimum for maximum callus production (Fig. 2). Present results support the study of Kasumi et al. (1998) (studying with Gladiolus cv. Topaz) in which similar result with same auxin concentration. Exogenous auxins have earlier been reported to induce somatic embryogenesis in callus as well as suspension cultures of Gladiolus (Kamo et al., 1990; Stefaniak, 1994; Remotti, 1995).
Upon to transfer to MS basal medium supplemented with 0.1 mg L-1 BA, creamy-white and friable calli produced numerous somatic embryos within 4 weeks (On avarage 30 structures per callus) (Fig. 3). There was not any embryo formation on other media. However presence of BA was necessary for normal development of somatic embryos (Endress, 1994), optimum BA concentration was required for embryo maturation may dependent on endogenous growth regulator level of explant.
 | |
| Fig. 1: | Shoots obtained from lateral buds on MS basal medium with 2 mg L-1 BA after 8 weeks from culture initiation, bar = 1 cm |
| Table 1: | Number and length of shoots in vitro obtained from lateral buds of Gladiolus anatolicus (Boiss.) Stapf cultured on MS medium supplemented with different BA concentrations |
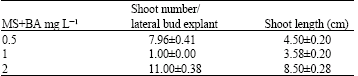 | |
| All experiments were conducted three replicates consisting of four explants per flask and were repeated twice. Data represented as mean±SE (standard error) from two experiments | |
| Table 2: | Effects of plant growth regulators on callus formation from the different regions of leaf in vitro obtained lateral bud explants of Gladiolus anatolicus (Boiss.) Stapf |
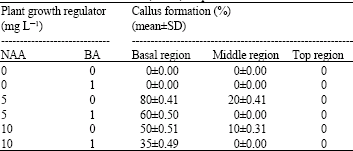 | |
| The experiments were conducted with three replicates consisting of four explants per flask and all experiments were repeated twice. The (%)±Standard deviation (SD) of two replicates. % mean±SD (standard deviation) | |
After 4 weeks, well-developed somatic embryos were transferred to MS basal medium supplemented with or without 0.1 mg L-1 BA and same media containing reduced sucrose concentration (20 g L-1 sucrose) for somatic embryo germination. The maturated somatic embryos were germinated easily upon transfer to MS basal medium supplemented with 0.1 mg L-1 BA but 20 g L-1 sucrose in two weeks (Fig. 4). On this medium 40% of the somatic embryos developed into plantlets.
 | |
| Fig. 2: | Creamy-white and friable calli on MS basal medium supplemented with 5 mg L-1 NAA, bar = 1 cm |
 | |
| Fig. 3: | Somatic embryos on MS basal medium supplemented with 0.1 mg L-1 BA, bar = 0.1 cm |
Kumar et al. (2002) working with Gladiolus hybridus Hort. were obtained the same result on MS medium with the same concentration of sucrose and without plant growth regulator.
The plantlets when transferred to MS basal medium (with 30 g L-1 sucrose) supplemented with 0.1 mg L-1 BA formed cormlets within 7-8 weeks (Fig. 5). Although Kumar et al. (2002) were reported cormlet formation was observed on MS basal media without growth regulators and supplemented with 80 g L-1 sucrose within 15 weeks, in this study, cormlet formation was only observed on MS basal media supplemented with 0.1 mg L-1 BA within 7-8 weeks.
 | |
| Fig. 4: | Germination of a somatic embryo on MS medium with 0.1 mg L-1 BA and 20 g L-1 sucrose, bar = 0.5 cm |
 | |
| Fig. 5: | Plantlets with cormlets on MS basal medium with 0.1 mg L-1 BA |
Optimum BA concentration is required for cormlet initiation may be dependent on endogenous growth regulator level.
To our knowledge this is the first report on somatic embryogenesis from leaf explant of Gladiolus anatolicus. We hope that will be helpful to breeding programs for large-scale clonal propagation of G. anatolicus.
REFERENCES
- Hussey, G., 1975. Totipotency in tissue explants and callus of some members of the Liliaceae, Iridaceae and Amaryllidaceae. J. Exp. Bot., 26: 253-262.
CrossRefDirect Link - Kamo, K., J. Chen and R. Lawson, 1990. The establishment of cell suspension cultures of Gladiolus that regenerate plants. In vitro Cell. Dev. Biol., 26: 425-430.
Direct Link - Kumar, A., A. Sood, L.M.S. Palni and A.K. Gupta, 1999. In vitro propagation of Gladiolus hybridus Hort. Synergistic effect of heat shock and sucrose on morphogenesis. Plant Cell Tissue Organ Cult., 57: 105-112.
Direct Link - Kumar, A., L.M.S. Palni, A. Sood, M. Sharma, U.T. Palni and A.K. Gupta, 2002. Heat-shock nduced somatic embryogenesis in callus cultures of gladiolus in the presence of high sucrose. J. Hortic. Sci. Biotechnol., 77: 73-78.
Direct Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Sen, J. and S. Sen, 1995. Two step bud culture technique for a high frequency regeneration of gladiolus corms. Sci. Hortic., 64: 133-138.
Direct Link - Stefaniak, B., 1994. Somatic embryogenesis and plant regeneration of Gladiolus (Gladiolus hort.). Plant Cell Rep., 13: 386-389.
Direct Link - Ghosh, B. and S. Sen, 1994. Plant regeneration from alginate encapsulated somatic embryos of Asparagus cooperi baker. Plant Cell Rep., 13: 381-385.
CrossRefDirect Link - Ziv, M., A.H. Halevy and R. Shilo, 1970. Organs and plantlets regeneration of Gladiolus through tissue culture. Ann. Bot., 34: 671-676.
CrossRefDirect Link