
M. Ranjbar
Department of Agronomy and Plant Breeding, Agricultural College, Tehran University, Karaj, Iran
M.R. Naghavi
Department of Agronomy and Plant Breeding, Agricultural College, Tehran University, Karaj, Iran
A. Zali
Department of Agronomy and Plant Breeding, Agricultural College, Tehran University, Karaj, Iran
M.J. Aghaei
Seed and Plant Improvement Institute, National Plant Gene Bank of Iran, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 7 | Page No.: 1126-1129







ABSTRACT
Abstract: Principal component and cluster analyses were used to evaluate the pattern of morphological variation in 122 accessions of Aegilops crassa for 14 quantitative characters. With the principal component analysis, the first five principal components with eigenvalues more than 1 contributed 69.5% of the variability amongst accessions, whereas PC6 to PC14 were less than unity. Plant height, stem diameter, spike length and number of spikelets per spike were the most important characters in the first principal component. The germplasm was grouped into five clusters using cluster analysis. Although each cluster had some specific characteristics of its own, but clusters were not clearly separated when plotted by the first two principal components. Mahalanobis distances (D2) determined that plant height, stem diameter, spike length, number of spikelets per spike, node width, seed length, seed width and flowering date characters as the most important characters in differentiating the accessions. The morphological variation of Aegilops crassa accessions obtained in this study provides useful information for the future collection and makes these genetic resources more accessible to breeders.
PDF Abstract XML References Citation
How to cite this article
M. Ranjbar, M.R. Naghavi, A. Zali and M.J. Aghaei, 2007. Multivariate Analysis of Morphological Variation in Accessions of Aegilops crassa from Iran. Pakistan Journal of Biological Sciences, 10: 1126-1129.
DOI: 10.3923/pjbs.2007.1126.1129
URL: https://scialert.net/abstract/?doi=pjbs.2007.1126.1129
DOI: 10.3923/pjbs.2007.1126.1129
URL: https://scialert.net/abstract/?doi=pjbs.2007.1126.1129
INTRODUCTION
Aegilops is mainly distributed in West Asia and the Mediterranean region and its center of diversity follows the central part of the Fertile Crescent arc in West Asia (van Slageren, 1994; Hegde et al., 2002). The genus Aegilops consists of approximately 21 species, classified into six sections or subgenera (Eig, 1929; van Slageren, 1994).
This genus has an important potential utilization in wheat improvement because of its resistance to different biotic and abiotic stresses and close relation with the cultivated wheat. Therefore, a better knowledge of the eco-geographical distribution of Aegilops species and their collection and conservation are required (Zaharieva et al., 2003). Low within-species genetic variation was observed in polyploid Aegilops species compared with diploid Aegilops species (Hegde et al., 2002).
Aegilops crassa BOISS. (tetraploid and hexaploid, genome symbols being DDMcrMcr and DDD2D2McrMcr, respectively) is known to be distributed through Palestine, Syria, Iraq, Iran, Afghanistan and Turkestan (Eig, 1929). It was reported previously by Kihara et al. (1957) that all the strains of Ae. crassa from Iran and Iraq are 4x, while 6x forms grow mixed together with 4x forms in Afghanistan and that the strains from Turkestan are 6x.
Phenotypic identification based a description of the morphological and physiological traits can be provided a simple way of quantifying genetic variation and have been successfully used for genetic diversity analyses and cultivar development (Smith and Smith, 1992). There are some research regarding morphological variation in Aegilops species (Zaharieva et al., 2003; Naghavi and Amirian, 2005; Knaggs et al., 2000; Leilah and Al-Khateeb, 2005). But still there are some regions that little or no information is available for important agronomic and morphological traits of these species. Although rich germplasm of Ae. crassa has been reported from Iran (Eig, 1929), but little information is available about the diversity of their accessions. The objective of this study was to investigate the morphological variation in a collection of Ae. crassa accessions sampled from different geographical regions of Iran.
MATERIALS AND METHODS
A total of 122 accessions of Aegilops crassa provided by the National Plant Gene Bank of Iran (NPGBI) were used for their diversity evaluation. Each accession was grown in a single row plot of 1 m long and wide and plant to plant spacing was 5 cm in a row, with three replications in a randomized complete block design.
Fourteen quantitative characters were recorded using descriptors established for Aegilops (IBPGR, 1981) with some modifications. Flowering date (days from sowing to appearance of 50% flowers) and maturity date (days from sowing to appearance of first maturity) were recorded by a single value for each row. Plant height (cm), stem diameter (mm), spike length (cm), spike diameter (mm), number of spikelets per spike, node length (cm), node width (mm), number of seeds per spikelet, culm length (cm), culm width (mm), seed length (cm) and seed width (mm) from 5 plants which had been randomly chosen in each row and mean of quantitative data sets were used for analysis.
The data recorded were analyzed for numerical taxonomic techniques using the procedure of principal component and cluster analyses (Hair et al., 1992) with the help of computer software Statistica and Spss. As the characters were recorded on different scales, the data were standardized to a mean of zero and a variance of unity prior to principal components analyses to eliminate scale differences (Sneath and Sokal, 1973). The principal components analysis is a multivariate statistical technique for exploration and simplifying complex data sets and has been demonstrated by Everitt and Dunn (1992).
Cluster analysis was conducted on the basis average distance of k-means and the accessions in each cluster were analyzed for basic statistics. The accessions of each of the five clusters were plotted according to their first two principal component scores. The relationships among the clusters were assessed by measuring the inter-cluster distances using Mahalanobis distance (D2).
RESULTS AND DISCUSSION
Principal component analysis was done to determine which of the characters more strongly contributed to the principal components. There are no tests to evaluate the significance of eigenvalues. Therefore, we follow the criterion established by Kaiser (1960), which adapts very well to the purpose of this analysis. This criterion is based on the selection of principal components whose eigenvalues are >1. Principal component analysis reduced the original 14 quantitative characters in experiment to 5 principal components. The first five principal components with eigenvalues >1 explained 69.5% of variation among 122 accessions (Table 1). Other Principal Components (PCS) had eigenvalues <1 and have not been interpreted. There are no clearly guidelines to determine the importance of a trait coefficient for each principal component. Johnson and Wichern (1988) regard a coefficient as significant that is greater than half divided by the square root of the standard deviation of the eigenvalue of the respective principal component.
| Table 1: | Principal components (PCS) for 14 characters in 122 accessions of Aegilops crassa |
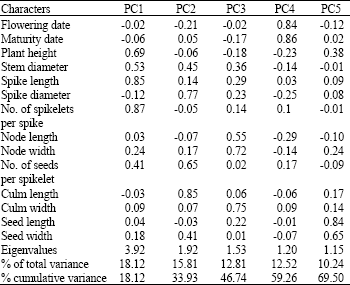 | |
But we used coefficient greater than half to decide the importance of characters in the different principal components. The first PC, which is the most important component, explained 18.12% of total variation and was positively related to plant height, stem diameter, spike length and number of spikelets per spike, so PC1 is a weighted average of these four characters. PC2 accounted of 15.81% of the total variation and the characters with the greatest weight on this component were spike diameter, number of seeds per spikelet and culm length. PC3 was mainly related to node length, node width and culm width, while PC4 was related to flowering date and maturity date and PC5 was related to seed length and seed width. Leilah and Al-Khateeb (2005) also found that first three principal components contributed 74.4% of the variability for 10 quantitative traits in wheat. Their results showed that PC1 correlated moderately well with plant height, number of spikes/m2, 100-grain weight and weight of grain/spike.
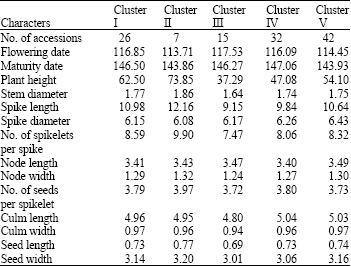
The entries in this study were grouped into five clusters based on average linkage, each cluster containing accessions that were highly similar. Cluster I consisted of 26 accessions, cluster II of 7, cluster III of 15, cluster IV of 32 and cluster V of 42 accessions. Mean value for each cluster (Table 2) revealed that accessions in cluster I characteristically had moderately late flowering and maturing and were low or medium for most characters, while accessions grouped in cluster II were tall and also the highest for stem diameter, spike length, number of spikelets per spike, node width, seed length and seed width. Accessions in cluster III and IV showed late flowering and maturity, respectively. Meanwhile accessions in cluster V were the highest for spike diameter and medium for other characters. The information on clusters to which particular accessions with traits of interest belong will assist in looking extensively for more accessions with similar traits (Upadhyaya et al., 2001)
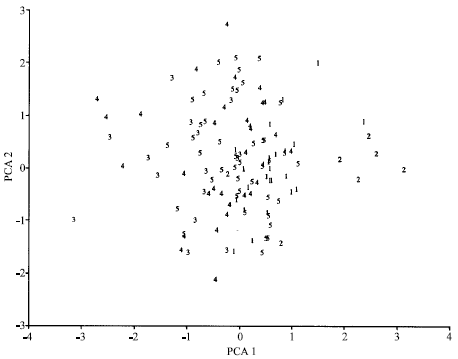 | |
| Fig. 1: | Principal component analysis for 14 characters in Iranian accessions of Aegilops crassa. 1, 2, 3, 4 and 5 represent the cluster number |
| Table 2: | Comparison profile of the five clusters of Aegilops crassa accessions classified by k-means clustering |
 | |
The accessions of each cluster and the first two principal component scores were plotted to aid visualization of group differences (Fig. 1). Clusters were not clearly separated which may be due to mixture of accessions with different morphological traits. Moreover accessions from same or near geographical origins scattered in different parts of the plot. In contrast, Ayana and Bekele (1999) grouped accessions of sorghum according to similar agro-climatic conditions and suggested the relevance of environmental factors in ordering the structure of variation in sorghum accessions.
| Table 3: | Pairwise Mahalanobis distances (D2) between five clusters of 122 accessions of Aegilops crassa |
 | |
According to the Mahalanobis distances (D2) among clusters (Table 3), the five clusters were statistically different from each other. The most similar clusters were, on the one hand, Clusters 4 and 5 (9.34 units) and, on the other, Clusters 1 and 5 (10.46 units). The largest inter-cluster distance (169.48 units) was between clusters 2 and 3. This result suggested that plant height, stem diameter, spike length, number of spikelets per spike, node width, seed length, seed width and flowering date are the most important characters in differentiating the accessions.
In our analysis, we were able to define groups of accessions that were significantly different from each other for characters of interest. Various multivariate methods, such as the principal components analysis and cluster analysis used here, have been shown to have utility in revealing germplasm grouping in numerous genetic resources such as blackgram (Ghaphor et al., 2001), sorghum (Ayana and Bekele, 1999; Grenier et al., 2001), quinoa (Ortiz et al., 1998), sugarcane (Tie and Miller, 2002) cacti (Carmona and Casas, 2005) and chickpea (Ghafoor et al., 2003; Naghavi and Jahansouz, 2005). In some of these studies suggested that morphological variation is strongly influenced by environmental factors. While in some others, like our study, both effects of environmental differences and genetic factors were important.
CONCLUSIONS
An understanding of the genetic variability available in germplasm collections is a preliminary requirement for the exploitation of useful traits in plant breeding. In this research we found that there was no relationship between genetic divergence and geographical origins as accessions from one place entered into more than one cluster. Conversely, accessions from different geographical origins were relatively unique and tend to be clustered in one part of the dendrogram. This information suggests that the morphological variation analyzed is determined not only by environmental differences but also by genetic factors. The lack of regional differentiation observed by the cluster and principal component analyses could be due to gene flow between regions. Anyway this high amount of morphological variation in Iranian accessions of Aegilops crassa makes these genetic resources more accessible for the future collection and provides these genetic resources more available to breeders.
REFERENCES
- Ayana, A. and E. Bekele, 1999. Multivariate analysis of morphological variation in sorghum (Sorghum bicolor (L.) Moench) germplasm from Ethiopia and Eritrea. Genet. Resour. Crop Evol., 46: 273-278.
CrossRefDirect Link - Carmona, A. and A. Casa, 2005. Management, phenotypic patterns and domestication of Polaskia chichipe (Cactaceae) in the Tehuacan valley, centeral Mexico. J. Arid Environ., 60: 115-132.
Direct Link - Ghaphor, A., A. Sharif, Z. Ahmad, M.A. Zahid and M.A. Rabbani, 2001. Genetic diversity in blackgram (Vigna mungo L. Hepper). Field Crop Res., 69: 183-190.
Direct Link - Grenier, C., P.J. Bramel-Cox and P. Hamon, 2001. Core collection of sorghum: I stratification based on eco-geographical data. Crop Sci., 41: 234-240.
Direct Link - Hegde, S.G., J. Valkoun and J.G. Waines, 2002. Genetic diversity in wild and weedy Aegilops, Amblyopyrum and Secale species-a preliminary survey. Crop Sci., 42: 608-614.
Direct Link - Kaiser, H.F., 1960. The application of electronic computers to factor analysis. Educ. Psychol. Measur., 20: 141-151.
CrossRef - Knaggs, P., M.J. Ambrose, S.M. Reader and T.E. Miller, 2000. Morphological characterisation and evaluation of the subdivision of Aegilops tauschii Coss. Wheat Inform. Service, 91: 15-19.
Direct Link - Leilah, A.A. and S.A. Al-Khateeb, 2005. Statistical analysis of wheat yield under drought conditions. J. Arid Environ., 61: 483-496.
Direct Link - Naghavi, M.R. and R. Amirian, 2005. Morphological characterization of accessions of Aegilops tauschii. Int. J. Agric. Biol., 7: 392-394.
Direct Link - Naghavi, M.R. and M.R. Jahansouz, 2005. Variation in agronomic and morphological traits among Iranian chickpea accessions. J. Integ. Plant Bio., 47: 375-379.
Direct Link - Ortiz, R., E.N. Ruiz-Tapia and A. Mujica-Sanchez, 1998. Sampling strategy for a core collection of Peruvian quinoa germplasm. Theor. Applied Genet., 96: 475-783.
Direct Link - Smith, J.S.C. and O.S. Smith, 1992. Fingerprinting crop varieties. Adv. Agron., 47: 85-140.
CrossRefDirect Link - Tie, P.Y.P. and J.D. Miller, 2002. Germplasm diversity among four sugarcane species for sugar composition. Crop Sci., 42: 958-964.
Direct Link - Upadhyaya, H.D. and R. Ortiz, 2001. A mini core subset for capturing and promoting utilization of chickpea genetic resource in crop improvement. Theor. Applied Genet., 102: 1292-1298.
Direct Link - Zaharieva, M., A. Dimov, P. Stankova, J. David and P. Monneveux, 2003. Morphological diversity and potential interest for wheat improvement of three Aegilops L. species from Bulgaria. Genet. Resour. Crop Evol., 50: 507-517.
CrossRefDirect Link