
R. Hajiboland
Plant Science Department, University of Tabriz, 51666 Tabriz, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 7 | Page No.: 1011-1019







ABSTRACT
From flora of Northwest Iran, two Graminaceous species including Secale montanum and Dactylis glomerata with different distribution and coverage on Ni-rich soils were selected for study of tolerance mechanisms to Ni concentrations up to 100 μ M in hydroponic culture medium. Shoot and root growth of D. glomerata was inhibited up to 42 and 69% in response to 100 μ M Ni, respectively, while growth reduction of S. montanum was as low as 20% in shoot and no growth inhibition of root was observed. Different growth response was not the result of different Ni accumulation, two studied species demonstrated a similar accumulation potential for Ni. A higher endogenous and Ni-induced malate accumulation did not cause a higher tolerance to Ni in D. glomerata. In contrast, high endogenous and Ni-induced accumulation of cysteine particularly in shoot was observed in Ni tolerant S. montanum. Distribution of Ni among various fractions of plant tissues studied by gel filtration chromatography suggested also an efficient chelation of excess Ni by low molecular weight fraction in S. montanum but not in D. glomerata. A higher re-translocation of Ni from mature into growing young leaves was one of the other causes of higher susceptibility of D. glomerata to excess Ni. Activity of peroxidase and catalase was much stronger inhibited in D. glomerata than S. montanum and activity of polyphenol oxidase was induced in response to Ni toxicity in roots of S. montanum. A possible role of this enzyme, involving in lignification, for higher Ni tolerance of S. montanum was discussed.
PDF Abstract XML References Citation
How to cite this article
R. Hajiboland, 2007. Uptake, Transport, Re-translocation and Chelation of Ni in One Tolerant and One Susceptible Graminaceous Species. Pakistan Journal of Biological Sciences, 10: 1011-1019.
DOI: 10.3923/pjbs.2007.1011.1019
URL: https://scialert.net/abstract/?doi=pjbs.2007.1011.1019
DOI: 10.3923/pjbs.2007.1011.1019
URL: https://scialert.net/abstract/?doi=pjbs.2007.1011.1019
INTRODUCTION
The presence of phytotoxic amounts of heavy metals in the soil can lead to toxicity symptoms and the inhibition of growth of most plants. Uptake, accumulation, distribution and detoxification of heavy metals, have been studied in a wide range of plants species (Clemens, 2001, Hall, 2002). However, the mechanisms involved are still only partially understood. The response of plants is complex with considerable variation between species. Several sequestration and detoxification strategies are known to occur, different effects are also observed with different metals and metal concentrations (Hall, 2002).
Complexation with phytochelatin peptides has been identified as an important mechanism for detoxifying metals such as Cd, but they have no apparent role in the detoxification of Zn and Ni (Cobbett, 2000).
Complexation of Ni by organic acids was studied in some Ni accumulator plants and it was shown that nickel is bound primarily to malic and malonic acids in Alyssum and to citric acid in many other hyperaccumulators such as Sebertia acuminata, Homalium and Hybanthus (Brooks, 1998a). However, from stability constants for complexes between nickel and amino acids and carboxylic acids it is clear that nickel complexes with amino acids are considerably more stable than those with carboxylic acids (Homer et al., 1997). Works of many investigators (Krämer et al., 1996) has displayed compelling evidence that nickel is bound to free histidine in Alyssum species. However, works on Ni-hyperaccumulator T. goesingense suggests that the histidine response might not be the general Ni-tolerance mechanism of plants (Persans et al., 1999).
An association between metals and low molecular weight organic compounds suggests that metal detoxification occurs by vacuolar sequestration (Boominathan and Doran, 2003). However, other strategies for metal tolerance and accumulation, such as binding to the cell wall or localization in the apoplast, may also be involved (Hall, 2002). Therefore, study of metal-organic and -amino acid complexes using chromatography (Brooks, 1988a) could provide information on the distribution of metals within various fractions of plant tissues as an indirect indicator of detoxification mechanism.
Uptake, transport and re-translocation of heavy metals could provide also additional information about the mechanisms of tolerance. Although the effect of nutrients re-translocation on the efficiency of plant genotypes was extensively studied (Marschner, 1995), the significance of re-translocation of elements to growing young leaves in the expressing of toxicity and/or differences among species was not studied.
Heavy metals are known to produce active oxygen species (Halliwell and Gutteridge, 1984). Activity of enzymes involving in the metabolism of free oxygen radicals e.g., peroxidase and catalase, was reported to be increased or decreased in response to metal toxicity depending on metal concentration, treatment duration or plant species (Chen et al., 2000; Sandalio et al., 2001; Schützendübel et al., 2001; Boominathan and Doran, 2002). Activity of polyphenoloxidase, involving in the biosynthesis of lignin, may be also influenced by metal toxicity as a consequence of common response of plants to stress. The extent of changes in the activity of these enzymes could affect the response of a given species to heavy metal toxicity.
Regarding contradictory results of chelation form and distribution of Ni between various cell fractions as well as effect of antioxidant capacity in determinations of plants response to heavy metal toxicity, it has been suggested that cellular mechanisms of heavy metal detoxification are highly species specific (Schat and Kalffl, 2000). In our previous study approximately 73 species were collected and identified from heavy-metal rich soils of Northwest of Iran (Hajiboland et al., 2002). Mechanisms of Ni tolerance of species growing on these soils are completely unknown. Moreover, some contrasting species concerning frequency and coverage on Ni-rich soils were introduced in this work (Hajiboland et al., 2002). From these contrasting species, Secale montanum and Dactylis glomerata both from Poaceae were of special interest and selected for further studies.
This research was aimed to study the mechanisms of Ni tolerance and/or susceptibility of two contrasting graminaceous species with special emphasize on chelation, fractionation in plants tissues as well as re-distribution of Ni among various organs in order to find the reason of different frequency and coverage of these two species on Ni-rich areas of Northwest of Iran.
MATERIALS AND METHODS
Plant materials: Two species were used in this study including Secale montanum and Dactylis glomerata. Both of plant species have been previously determined as members of flora of Ni rich soils in NW Iran. Seeds were collected from Ni rich areas near Khoy (West Azarbaijan Province, Iran).
Plants culture: The experiments were conducted in a growth chamber under controlled environmental conditions. Seven-days-old seedlings were transferred to the nutrient solution (Cakmak et al., 1996) and pre-cultured for 3 days. Nickel treatments were started for 10-days-old plants, consisted of five levels of NiSO4 at 0 (control), 25, 50, 75 and 100 μM.
Harvest: After 12 days treatment, plants were harvested. For removing of the apoplasmic Ni from roots, plants were placed for 1 h in 5 mM CaCl2+25% nutrient solution without Ni. Thereafter, roots were washed with distilled water, divided into shoots and roots, weighed and blotted dry on filter paper and dried at 70°C for 2 days to determine plant dry weight. Nickel concentration was determined by atomic absorption spectrophotometry (Shimadzu, AA 6500).
TLC: Fresh tissue samples were extracted in 20 mM Tris-HCl (pH = 8.0). The crude extract was centrifuged and applied to the plates according to the method described by Oven et al. (2002). Tissue extract was applied to a cellulose plate (formic acid/ethanol/water 48.8:48.8:2.4, v/v/v as mobile phase) for organic acids and to a silica gel plate (isobuthanol/acetic acid/water 50:25:25, v/v/v as mobile phase) for amino acids. After drying for 1 day at room temperature, dichlorophenol-indophenol and ninhydrin reagents were used for detection of organic and amino acids, respectively. The standard solutions of citric acid, malic acid, histidine, cysteine, glutamine and asparagines were applied simultaneously to the plates.
After detection of spots using an UV-detector, the margins of spots were lined and presence or absence as well as relative amount of each compound were reported using-or+characters. Each TLC experiment was conducted in triplicate.
Cysteine assay: Extraction of cysteine was carried out according to the method was described for TLC method. A quantitative determination of cysteine in plant tissues was conducted using a photometric method (Gaitonde, 1967).
Size exclusion chromatography: The fractionation of tissue extract of plants treated for 1 week with 50 μ M Ni, was carried out according to the method described by Guo (1995). Fresh shoot or root tissues were homogenized in 10 mM Tris-HCl (pH = 8.0, +100 mM NaCl) in a chilled mortar then centrifuged at 2000 xg for 10 min at 4° C. The liquid fraction was re-centrifuged at 30000 x g for 45 min at 4° C. The supernatant (soluble fraction) was applied on a column of Sephadex G-50 fine (50x 2.5 cm) pre-equilibrated with running buffer and eluted at a flow rate of 60 mL h- 1. The running and pre-equilibration buffer was the same and contained 10 mM Tris-HCl (pH = 8.0) supplemented by 100 mM NaCl. Elutant solution was collected in 60x 1 mL samples. The concentration of Ni in each fraction was determined by AAS.
Re-translocation experiment: The experiment was carried out using method described previously for Cu (Hajiboland and Boiadi, 2005). The first harvest was performed at the end of root washing and four subsequent harvests were carried out each time after that a new leaf had emerged and partially expanded. Four replicates, consisting of defined leaves of four plants, were harvested for each treatment and harvest time. Samples were washed with double distilled water, oven-dried samples were ashed and Ni concentration was determined by AAS.
Assay of enzymes activity: Catalase (EC 1.11.1.6) activity was assayed by monitoring the decrease in absorbance of H2O2 at 240 nm (Lück, 1962). Unit activity was taken as the amount of enzyme, which decomposes 1 M of H2O2 in one min. Peroxidase (EC 1.11.1.7) activity was determined using the guaiacol test (Chance and Maehly, 1955), the absorption of tetraguaiacol formed in the reaction was measured at 470 mm. The enzyme unit was calculated as enzyme protein required for the formation of 1 μ M tetraguaiacol for 1 min. Polyphenoloxidase (EC 1.10.31) activity was assayed by monitoring the absorbance at 430 nm (Halder et al., 1998). The enzyme unit was calculated as enzyme protein required for color change for 1 min.
Determination of protein in tissue extracts for calculation of enzymes activities was carried out according to Bradford (1976).
RESULTS
Plants growth and accumulation of Ni in shoots and roots: Shoot and root dry weight of both species decreased in response to toxic levels of Nickel. However, growth reduction of D. glomerata was much higher than that of S. montanum. Shoot and root growth of D. glomerata was inhibited up to 42 and 69% in response to 100 μM Ni, respectively. The growth inhibition in S. montanum was only 20% in shoot. In roots no significant growth inhibition was observed at Ni treatment as high as 100 μM (Table 1).
Concentration of Ni in roots of plants grown at high Ni levels in medium was much higher than shoots. On the other hand, the extent of Ni accumulation was similar in two studied species (Table 1).
Accumulation of organic acids, amino acids and amides: In shoot extract of S. montanum organic acids were not detectable with TLC method. But in shoot extract of D. glomerata not only malate and citrate were detected, but also an increase in malic acid content in plants treated by 50 μ M Ni was observed. In root extract of both species detectable amounts of malate and citrate was found, but similar with shoot, the increase in malic acid content was observed only in D. glomerata and no detectable change in citric acid amounts was observed in both species (Table 2).
| Table 1: | Growth and concentration of Ni in shoot and roots of Dactylis glomerata and Secale montanum grown at toxic levels of this metal in the nutrient solution |
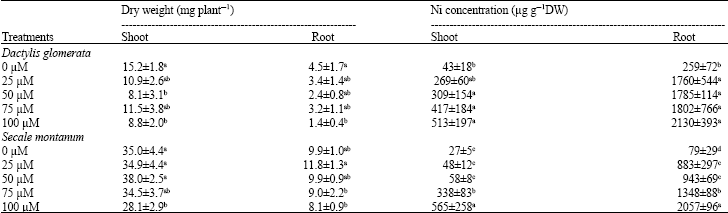 | |
| Values in each column within each plant species followed by the same letter(s) are not significantly different (p<0.05) | |
| Table 2: | Detection of the presence of organic acids by TLC in tissue extract of Dactylis glomerata and Secale montanum grown at toxic levels of Ni in the nutrient solution |
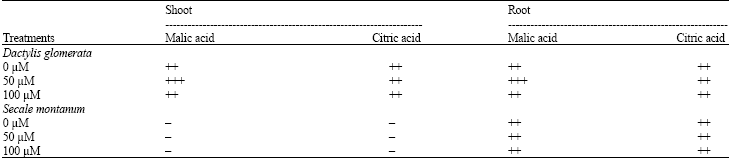 | |
| The number of +signs is correlated with the concentration | |
Histidine was not dateable in both shoot and root extracts and in both studied species neither in control nor in Ni treated plants. Cysteine was found in prominent amounts and in both plant organs. An increase in cysteine amount in response to Ni treatment was observed in shoot of S. montanum. Glutamine but not asparagine was detected in both shoot and root extract of D. glomerata, but not of S. montanum. However, no change in response to Ni treatment was observed in glutamine content of shoot and root (Table 3).
Data of cysteine concentration which were determined spectrophotometrically, were in accordance with TLC experiment, so that a significant increase in cysteine concentration of shoot in response to 50 μM Ni was observed in S. montanum. In D. glomerata cysteine concentration of shoot rather decreased in response to 50 μM Ni. In roots of D. glomerata a significant reduction and in S. montanum an increase in cysteine concentration was observed in response to Ni (Table 4). Changes in cysteine concentration of root in response to Ni treatment could not be detected by TLC method.
Distribution of Ni between various fractions of shoot and root: To characterize the binding state of Ni in shoot and roots, the soluble fraction was eluted through a Sephadex column. In the soluble fraction, phytochelatins can be separated from proteins and Low-Molecular-Weight (LMW) solutes by gel filtration (Mutoh and Hayashi, 1991). Accordingly, the soluble fraction of shoot and root extracts was separated into 60 fractions. The concentrations of Ni in these fractions are presented in Fig. 1. The first peak between fraction 18 and 22 represents High Molecular Weight Compounds (HMWs), the second peak between fraction 28 and 41 most likely contains PCS (Becher and Höfner, 1994) and the third peak between fraction 45 and 59 corresponds to LMW, such as organic acids, amino acids and probably also free metal ions (Mutoh and Hayashi, 1991). In the chromatogram (Fig. 1), it could be detected two peaks in shoot and one peak in root extract. The peaks of root extract were similar in two studied species and an exact coincidence was not observed between these two chromatograms with given data by other workers for elution time of three mentioned group of compounds. These peaks could represent the MMWs (phytochelatins) or may represent the LMWs, organic and amino acids as well as free Ni.
The chromatogram of the shoot extract was different between two species. On the other hand, according to elution time, it is possible to determine with a high confidence the nature of compounds to them Ni was chelated. In D. glomerata, the first peak was coincided on the HMWs, possibly proteins and other soluble macromolecules and the second peak represented the MMWs, possibly phytochelatins and other small peptides. In S. montanum, the first peak was coincided on the MMWs, possibly phytochelatins and the second peak represented the LMWs, organic and amino acids as well as free Ni. In LMWs fraction, Ni could not be detected in D. glomerata and in HMWs fraction in S. montanum (Fig. 1).
| Table 3: | Detection of the presence of amino acids by TLC in tissue extract of Dactylis glomerata and Secale montanum grown at toxic levels of Ni in the nutrient solution. |
 | |
| The number of +signs is correlated with the concentration | |
| Table 4: | The concentration of cysteine ( μ g g- 1FW) detected spectrophotometrically in tissue extract of Dactylis glomerata and Secale montanum grown at toxic levels of Ni in the nutrient solution |
 | |
| Values in each column followed by the same letter(s) are not significantly different (p<0.05) | |
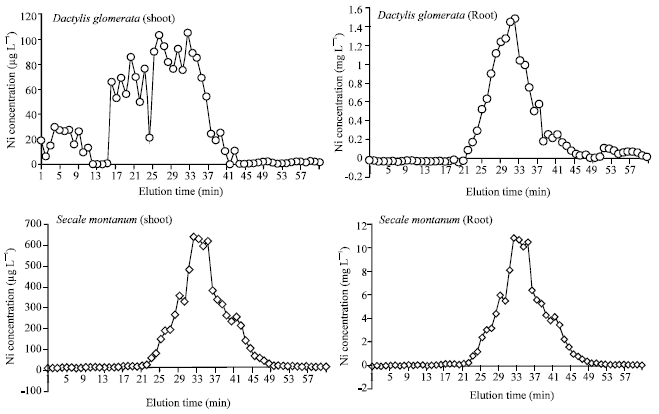 | |
| Fig. 1: | Concentration of Ni in the eluted fractions of shoot ( μ g L- 1) and root (mg L- 1) extracts from gel-chromatography on a Sephadex G50 column in D. glomerata and S. montanum grown at 50 μ M Ni in medium |
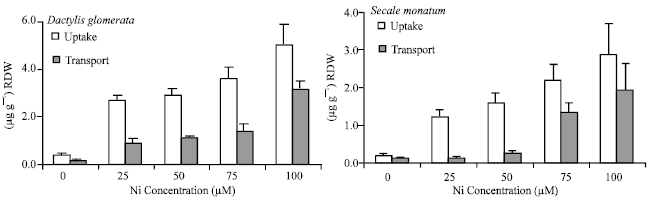 | |
| Fig. 2: | Uptake ( μ g g- 1 RDW) and transport ( μ g g- 1 RDW) of Ni in D. glomerata and S. montanum at toxic concentrations of Ni in medium |
Uptake, transport and Re-translocation of Ni from mature to young leaves: As expected, uptake and transport of Ni increased with increasing concentration of metal in the medium. However, transport into the shoot increased more distinctly at higher Ni treatments, particularly in S. montanum. The Ni transport into shoot amounted only 14% of total Ni taken up at Ni concentration of 50 μ M, while up to 67% of total Ni in plants treated with 100 μ M Ni, was transported into aerial part (Fig. 2).
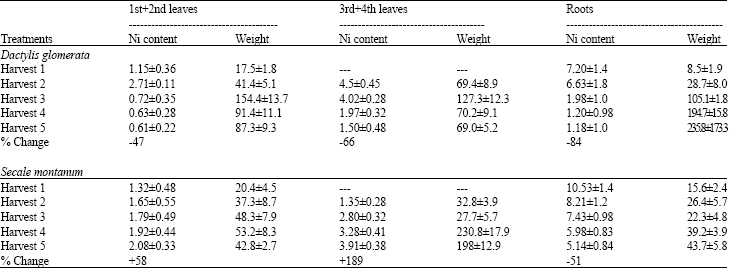
Ni content of each distinct leaves and roots changed significantly during the experimental period as simultaneous increase of leaf and root weight occurs. Ni content of roots decreased continuously toward the end of experimental period. This reduction was higher in D. glomerata than S. montanum. In various fractions of shoots, however, more distinct difference between two studied species was observed. Nickel content of mature leaves decreased in D. glomerata but increased in S. montanum.
| Table 5: | Changes in the content ( μ g plant part- 1) and weight (mg plant part- 1) of various shoot fractions of Dactylis glomerata and Secale montanum loaded by Ni concentration of 100 μ M in the nutrient solution for 2 days and followed during various harvest intervals |
 | |
| Table 6: | Activity of peroxidase, catalase and polyphenoloxidase (unit mg- 1 Pro) in tissue extract of Dactylis glomerata and Secale montanum grown at toxic levels of Ni in the nutrient solution |
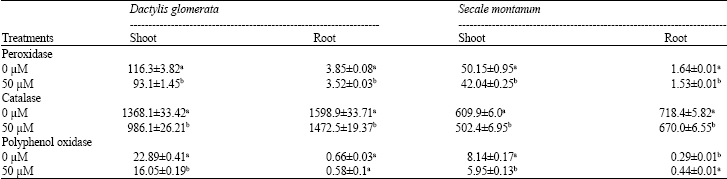 | |
| Values in each column within each enzyme activity followed by the same letter(s) are not significantly different (p<0.05) | |
It means that in D. glomerata after appearance of new growing young leaves, each mature leaf lost significant amounts of Ni loaded previously in these leaves and obviously re-translocate it to young leaf. In S. montanum such a reduction of Ni content of mature leaves did not occur and rather an increase in Ni content of old leaves was observed (Table 5).
Changes in the activity of enzymes in response to Ni treatment: Activity of three studied enzymes decreased in response to Ni treatment in both shoot and root. The extent of reduction was higher in shoots than roots in both species. In shoots % reduction of enzyme activities were higher in D. glomerata than S. montanum. The % reduction of peroxidase, catalase and polyphenoloxidase activity were 20%, 28 and 30% in D. glomerata but 16%, 18 and 27% in S. montanum. In roots the inhibition of enzyme activity in two studied species was similar. The activity of polyphenoloxidase was induced in roots of S. montanum in response to treatment with 50 μM Ni (Table 6).
DISCUSSION
A clear different growth response to excess Ni was observed between D. glomerata and S. montanum. This is in accordance with our observation on Ni rich soils. S. montanum is one of the important elements of Ni flora in NW Iran (Hajiboland, 2002). In contrast, the distribution of D. glomerata is mainly restricted to marginal areas of Ni soils with low Ni availability. A high tolerance to Zn toxicity was observed in S. montanum than D. glomerata in our previous work (Hajiboland et al., 2006).
The main cause of difference between two studied species was not different uptake and accumulation of Ni. Both species took up Ni and accumulated it in shoot with similar extent. Hyperaccumulators of Ni have a concentration of Ni in aerial parts as high as 1000 μg g-1 DW (Brooks, 1998b). In this research, the accumulation of Ni in leaves was 500 μg g-1 DW, therefore, these species do not quite qualify for hyperaccumulator status. The amounts of Ni concentration of plant samples collected from Ni soils, was in the range of 50-100 μg g DW.
Therefore, these two species could be classified only as accumulator species but with different tolerance to Ni toxicity.
Carboxylic acids and amino acids such as citric, malic and histidine are potential ligands for heavy metals and so could play a role in tolerance and detoxification (Rauser, 1999; Clemens, 2001), however, strong evidence for a function in tolerance, such as a clear correlation between amounts of acid produced and exposure to a metal, has not been produced to support a widespread role (Hall, 2002).
A clear difference between two species was observed in malate accumulation and induction in response to Ni in this work. However, a high endogenous and Ni-induced malate concentration (at 50 μM Ni) in D. glomerata was not associated with higher tolerance, suggesting that malate had no effect on tolerance of this species. The effect of malate as a Ni chelating and detoxifying molecule was reported for Psychotria douarrei (Kersten et al., 1980), but Ni chelation by malate was not observed in Sebertia acuminate, Homalium and Hybanthus (Brooks, 1988a). Malate is one of the important metabolites in TCA cycle and fluctuation of its concentration could be the result of a wide range of factors affecting the activity of TCA cycle enzymes.
In contrast to reports on Ni chelation by histidine in Alyssum lesbiacum (Krämer et al., 1996), in this study detectable amount of this amino acid was not found by TLC method, therefore, further study on histidine concentration was not carried out. It was also reported that, histidine response is not involved in Ni tolerance of all Ni hyperaccumulators (Persans et al., 1999). In contrast to histidine, a clear cysteine response was observed in Ni treated plants. Both TLC and spectrophotometry methods showed that, cysteine accumulated in response to Ni and induction of cysteine was associated with higher Ni tolerance in S. montanum. Apart from inductive effect of Ni, the absolute amounts of cysteine were significantly higher in S. montanum which could be an indication of a higher potential for tolerance to heavy metals in this species. For cobalt, one of the heavy metals that do not activate PC synthesis, the cysteine and citrate concentrations are elevated in the hyperaccumulator Crotalaria cobalticola (Oven et al., 2002). Our previous work showed also the effect of high endogenous and Cu-induced cysteine content of shoot in Cu tolerance of Astragalus echinops, a Cu tolerant species growing around Cu mines of Northwest of Iran (Hajiboland et al., 2006).
In shoots a clear difference in distribution of Ni among various parts of soluble fraction between two studied species was observed. A high Ni concentration was found in LMW fraction of shoot in S. montanum, most likely bound with cysteine as chelating molecule in this fraction. Ni concentration in LMW fraction of shoot extract of D. glomerata was negligible, therefore, a high sensitivity of D. glomerata to Ni toxicity, could be attributed to the lack of Ni in this fraction.
Considerable amounts of Ni in D. glomerata were found in HMWs, implicating the association of Ni with. proteins and structural macromolecules instead of chelation with low molecular weight compounds such as cysteine.
The presence of Ni in the MMWs, was similar between two species. This fraction may contain phytochelatins (Becher and Höfner, 1994). Although phytochelatin-Ni complex by phytochemical methods has not been detected (Cobbett, 2000), chelation of Ni to phytochelatins in these species could not be ruled out. However, similar portion of Ni in MMWs between two species suggested that chelation of Ni with compounds in this fraction has obviously no distinct role in detoxification likely because of a poor compartmentation.
This is the second report on the re-translocation of a heavy metal at toxic levels in a long term experiment using root-fed plants. Results suggested that the distribution of Ni among leaves of various age i.e. different effect on growth response, could be important than total Ni content. A high Cu re-translocation resulting in a higher susceptibility to excess Cu was reported for maize and sunflower (Hajiboland and Boiadi, 2005). The results for the Ni re-translocation showed also that, substantial quantities of Ni, loaded in roots, were transported into the shoots shortly after loading. The extent of root-shoot translocation (reflected in %reduction of Ni content of root) was higher in D. glomerata compared to S. montanum. On the other hand, in D. glomerata after 2-3 harvest intervals (18-27 days growth), the mature leaves became source of Ni and began to re-translocate the loaded Ni to younger leaves which was well reflected in the reducing amounts of Ni in mature leaves during the 3rd and 5th harvests in this species. In contrast, in S. montanum Ni content of mature leaves was not reduced during the experimental period. Therefore, the mature leaves of S. montanum did not act as a source of Ni for young leaves and simultaneous, acted as a sink for Ni loaded previously in roots.
Reduction in the activity of antioxidant enzymes e.g., peroxidase and catalase, was reported under toxicity of heavy metals including Ni (Boomanathan and Doran, 2002). Higher reduction of the activity of antioxidant enzymes in D. glomerata than S. montanum under Ni toxicity leading to oxidative stress, could be one of the other factors determining responses of two tested species to Ni toxicity. However, the activity of polyphenol oxidase in roots was not either influenced by Ni toxicity (D. glomerata) or rather increased (S. montanum). Polyphenol oxidase is involved in lignification of cell walls. Higher activity of polyphenol oxidase in roots of S. montanum in the presence of excess Ni could be one of the reasons for higher tolerance of this species particularly regarding to root growth. The heavy metal induced membrane damages and leakage of solutes from roots is well documented (Murphy et al., 1999). Therefore, induction of lignification could prevent leakage of solutes from roots. It was reported that, Cd (Schützendübel et al., 2001) and Cu (Chen et al., 2000; Díaz et al., 2001; Jouili and Ferjani, 2003) toxicity induce xylogenesis. In this study, a higher polyphenol oxidase activity likely leads to an induction of lignification and thereby providing a mechanism for adaptation against higher Ni concentrations for S. montanum. A possible role for lignification in establishing an apoplastic transport barrier in roots was hypothesized under Cu and Zn toxicity (Cuypers et al., 2002).
CONCLUSION
Different sensitivity to Ni toxicity between D. glomerata and S. montanum grown in hydroponic medium was in accordance with our ecological observation in terms of frequency and coverage of these species on Ni-rich areas in Northwest of Iran. Study of physiological mechanisms involving in their contrasting response to excess Ni revealed that, although both species have similar uptake and accumulation of Ni, different re-translocation potential for Ni in association with a different sensitivity at tissue level due to different extent of Ni chelation are from the important reasons of contrasting responses of these species to excess Ni.
ACKNOWLEDGMENTS
The author is grateful to the Research Council of Tabriz University for providing financial assistance (Project No. 16/359).
REFERENCES
- Becher, M. and W. Hofner, 1994. Demonstration of Cd-complexing compound in Scenedesmus subspicatus and Zea mays L. similar to phytochelatins. Z. Pflanzenernahr. Bodenk., 157: 87-92.
Direct Link - Boominathan, R. and P.M. Doran, 2002. Ni-induced oxidative stress in roots of the Ni hyperaccumulator, Alyssum bertolonii. New Phytol., 156: 205-215.
Direct Link - Boominathan, R. and P.M. Doran, 2003. Organic acid complexation, heavy metal distribution and the effect of ATPase inhibition in hairy roots of hyperaccumulator plant species. J. Biotechnol., 101: 131-146.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Cakmak, I., N. Sari, H. Marschner, M. Kalayci A. Yilmaz, S. Eker and K.Y. Gulut, 1996. Dry matter production and distribution of zinc in bread and durum wheat genotypes differing in zinc efficiency. Plant Soil, 180: 173-181.
Direct Link - Chance, B. and A.C. Maehly, 1955. Assay of catalases and peroxidases: Catalase: 2H2O2 → 2H2O + O2 (1), Catalase and peroxidase: ROOH + AH2 → H2O + ROH + A (2). Methods Enzymol., 2: 764-775.
CrossRefDirect Link - Chen, L.M., C.C. Lin and C.H. Kao, 2000. Copper toxicity in rice seedlings: Changes in antioxidative enzyme activities, H2O2 level and cell wall peroxidase activity in roots. Bot. Bull. Acad. Sin., 41: 99-103.
Direct Link - Clemens, S., 2001. Molecular mechanisms of plant metal tolerance and homeostasis. Planta, 212: 475-486.
CrossRefDirect Link - Cobbett, C.S., 2000. Phytochelatins and their roles in heavy metal detoxification. Plant Physiol., 123: 825-832.
CrossRefDirect Link - Cuypers, A.N.N., J. Vangronsveld and H. Clijsters, 2002. Peroxidase in roots and primary leaves of Phaseolus vulgaris under copper and zinc phytotoxicity: A comparison. J. Plant Physiol., 159: 869-876.
Direct Link - Diaz, J., A. Benal, F. Pomar and F. Merino, 2001. Induction of shikimate dehydrogenase and peroxidase in pepper (Capsicum annuum L.) seedlings in response to copper stress and its relation to lignification. Plant Sci., 161: 179-188.
CrossRefDirect Link - Gaitonde, M.K., 1967. A spectrophotometric method for the direct determination of cysteine in the presence of other naturally occurring amino acids. Biochem. J., 104: 627-633.
Direct Link - Hajiboland, R. and H. Boniadi, 2005. Accumulation of cu in root apoplasm and re-translocation to young leaves in rice, maize and sunflower with different toxicity tolerance. Pak. J. Biol. Sci., 8: 1599-1609.
CrossRefDirect Link - Halder, J., P. Tamuli and A.N. Bhaduri, 1998. Isolation and characterization of polyphenol oxidase from Indian tea leaf (Camelia sinensis). J. Nutr. Biochem., 8: 75-80.
Direct Link - Hall, J.L., 2002. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot., 53: 1-11.
CrossRefDirect Link - Halliwell, B. and J.M. Gutteridge, 1984. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J., 219: 1-14.
PubMed - Jouili, H. and E. El Ferjani, 2003. Changes in antioxidant and lignifying enzyme activities in sunflower roots (Helianthus annuus L.) stressed with copper excess. C.R. Biologies, 326: 639-644.
CrossRefDirect Link - Kersten, W.J., R.R. Books, R.D. Reeves and T. Jaffre, 1980. Nature of Nickel complexes in Psychotria douarrei and other Nickel accumulating plants. Phytochemistry, 19: 1963-1965.
Direct Link - Kramer, U., J.D. Cotter-Howells, J.M. Charnock, A.J.M. Baker and J.A.C. Smith, 1996. Free histidine as a metal chelator in plants that accumulate nickel. Nature, 379: 635-638.
Direct Link - Murphy, A.S., W.R. Eisinger, J.E. Shaff, L.V. Kochian and L. Taiz, 1999. Early vopper-induced leakage of K+ from Arabidopsis seedlings is mediated by ion channels and coupled to citrate efflux. Plant Physiol., 121: 1375-1382.
Direct Link - Mutoh, N. and Y. Hayashi, 1991. Sulfur-containing cadystin-cadmium complexes. Methods Enzymol., 205: 341-347.
Direct Link - Oven, M., E. Grill, A.G. Goldhirsh, T.M. Kutchan and M.H. Zenk, 2002. Increase of free cysteine and citric acid in plant cells exposed to cobalt ions. Phytochemistry, 60: 467-474.
Direct Link - Persans, M.W., X. Yan, J.M.L. Patnoe, U. Kramer and D. Salt, 1999. E Molecular dissection of the role of histidine in Nickel hyper accumulation in Thlapsi goesingense (Hapacsy). Plant Physiol., 121: 1117-1126.
Direct Link - Rauser, W.E., 1999. Structure and function of metal chelators produced by plants. Cell Biochem. Biophys., 31: 19-48.
CrossRef - Sandalio, L.M., H.C. Dalurzo, M. Gomez, M.C. Romero-Puertas and L.A. del Rio, 2001. Cadmium-induced changes in the growth and oxidative metabolism of pea plants. J. Exp. Bot., 52: 2115-2126.
Direct Link - Schat, H. and M.M.A. Kalff, 2000. Are Phytochelatines Involved in Different Metal Tolerance, Uptake and transport of Heavy Metals in Hyperaccumulating and non Hyperaccumulating Metallophytes. In: Phytoremediation of Contaminated Soil and Water, Terry, N. and G. Banuelos (Eds.). CRC Press, Boca Raton, FL., USA., pp: 171-188.
- Schutzendubel, A., P. Schwanz, T. Teichmann, K. Gross, R. Langenfeld-Heyser, D.L. Godbold and A. Polle, 2001. Cadmium-induced changes in antioxidative systems, hydrogen peroxide content and differentiation in scots pine roots. Plant Physiol., 127: 887-898.
CrossRefDirect Link