
Sadeghi, A.
Department of Genomics, Agricultural Biotechnology Research Institute of Iran, Mahdasht Road, Karaj, Iran
A.R. Hessan
Plant Diseases Lab., Plant Pests and Diseases Research Institute, Mahdasht Road, Karaj, Iran
H. Askari
Department of Plant Physiology, Biochemistry and Proteomics, Agricultural Biotechnology Research Institute of Iran
S. Aghighi
Departments of Plant Protection and Biotechnology, College of Agriculture, Shahid Bahonar University, 22 Bahman Blvd, Kerman, Iran
G.H. Shahidi Bonjar
Departments of Plant Protection and Biotechnology, College of Agriculture, Shahid Bahonar University, 22 Bahman Blvd, Kerman, Iran
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 5 | Page No.: 904-910







ABSTRACT
Biological control of sugar beet damping-off of Rhizoctonia solani by two Streptomyces isolates (S2 and C) was evaluated in this study. The in vitro antagonism assays showed that active isolates had inhibitory effects on mycelium growth of the three R. solani AG-4 isolates (Rs1, Rs2 and Rs3). Soil treatment either with isolate S2 or C formulation inhibited the disease completely and increased seedling stand in infected and uninfected treatments significantly (p<0.05). Compared to controls, all treatments containing bacteria had enhanced shoot and root dry weight and root density. Both bacterial isolates maintained normal growth in pH ranges of 5.6, 7.2 and 8.0 at 29°C. Isolate C grew in pH ranges 5.6, 7.2 and 8.0 at 23-37°C, isolate S2 grew at 18-37°C in pH 5.6-8.0 but did not grow in acidic medium at 37°C. When pH decreased to 5.0, growth of C decreased and S2 inhibited. To elucidate the mode of antagonism, chitinase activity and siderophore production were evaluated. Both isolates showed chitinase activity on medium containing colloidal chitin. Biosynthesis of siderophore was detectable in isolate C but not in S2. The results of this study showed that these isolates had antifungal activities by production of siderophore and chitinase.
PDF Abstract XML References Citation
How to cite this article
Sadeghi, A., A.R. Hessan, H. Askari, S. Aghighi and G.H. Shahidi Bonjar, 2006. Biological Control Potential of Two Streptomyces Isolates on Rhizoctonia solani, the Causal Agent of Damping-off of Sugar Beet. Pakistan Journal of Biological Sciences, 9: 904-910.
DOI: 10.3923/pjbs.2006.904.910
URL: https://scialert.net/abstract/?doi=pjbs.2006.904.910
DOI: 10.3923/pjbs.2006.904.910
URL: https://scialert.net/abstract/?doi=pjbs.2006.904.910
INTRODUCTION
Damping-off is a common problem in almost all field and greenhouse crops (Georgakopoulos et al., 2002). Rhizoctonia solani Kuhn is one of soilborne pathogenic fungi have been associated with damping off (Moussa, 2002). Protection of sugar beet against this pathogen is important to maximize the crop yield. The use of fungicide-treated seeds is the main commercially option for control of damping off. Development of biocontrol systems has provided an effective approach for reducing environmental pollution associated with fungicides and the risk of fungicide resistance development. Biological control of soil borne diseases by microorganisms especially Streptomyces species have already been published (Crawford et al., 1993; EL-Abyad et al., 1993; Asaka and Shoda, 1996; Jones and Samac, 1996; Chamberlain and Crawford, 1999; Whipps, 2001; Xiao et al., 2002).
Several species of Streptomyces have inhibitory effects on the most common soil borne fungi like Fusarium oxysporum (Smith et al., 1990; Abd-Allah, 2001; Getha and Vikineswary, 2002), Pythium ultimum (Crawford et al., 1993; Yuan and Crawford, 1995; Paulitz and Belanger, 2001), Verticillium spp. (EL–Abyad et al., 1993; Aghighi et al., 2004), Rhizoctonia solani (Asaka and Shoda, 1996; Sabaratnam and Traquair, 2002), Sclerotinia homeocarpa, Gaeumannomyces graminis and Microdochium nivale (Chamberlain and Crawford, 1999). According to recent publications, application of Streptomyces, (Sabaratnam and Traquair, 2002; Chung et al., 2005) or their active ingredient from the culture filtrate (Shih et al., 2003) are effective for control of Rhizoctonia damping off. Beside, some species are plant growth promoting agents that it may be due to the ability of these bacteria to produce hydroxamate-type siderophores and/or other plant growth-promoting metabolites (Nassar et al., 2003).
The mode of Streptomyces action includes inhibition of the pathogen by antifungal antibiotics (Smith et al., 1990; Gupte and Naik, 1999) competition for iron through production of siderophore; production of degradative enzymes such as chitinase and glucanase (El-Tarabily et al., 2000) and chitin binding proteins (Bormann et al., 1999). Trejo-Estrada (1998) reported antagonistic effects of a Streptomyces violaceusniger isolate on plant pathogens. They showed that biocontrol activity of this bacterium was resulted from three antifungal compounds; also the isolate was able to produce chitinase and glucanase. Results of De Boer (1998) revealed that antibiotics were involved in antifungal activities of chitinolytic bacteria. El-Tarabily et al. (2000) determined that production of chitinase and glucanase was the main mechanisms related to biocontrol activity of Streptomyces viridodiasticus. Besides, other compounds such as phenylacetic and sodium phenylacetate from Streptomyces humidus showed antifungal activity on plant pathogens (Hwang et al., 2001). Practical application of some beneficial biocontrol agents such as Pseudomonads has been limited due to decreased bacterial survival during commercial processing and packaging of formulation or after application in the field (Walker, 2004). In contrast, Streptomyces are able to produce spores to maintain their viability at non-specific conditions (Ensign, 1978). However, Yuan and Crawford (1995) showed that the density of the viable Streptomyces spores in formulation was only 10 fold less than origin over a 6.5-month period of storage at room temperature. Other significant parameter influencing the survival of bacteria in soil is temperature. Survival and antifungal activity (antibiotic and siderophore production) of Pseudomonads strains decrease at temperatures above 24°C (Schmidt et al., 2004) but Streptomyces are mesophilic bacteria and grow at a wide range of temperature between 15 and 37°C (Locci, 1994). These beneficial characteristics of Streptomyces have attracted many investigators for isolating and improvement of these bacteria in biocontrol programs.
In our preliminary study, in vitro inhibitory effects of two isolates of Streptomyces (S2 and C) were assessed against some important pathogenic fungi including Fusarium graminearum, F. oxysporum f. sp. melonis, Pyricularia grisea and Rhizoctonia solani which cause rice root rot, wheat head blight, melon damping- off, rice blast and rice sheath blight, respectively. Our main objectives in the present study were introduction of potentially active biocontrol Streptomyces isolates against R. solani, the causal agent of damping-off in sugar beets and partial characterization of their effectiveness on sugar beet growth. We also surveyed growth sensitivity of the Streptomyces isolates to temperature and pH ranges. To assess the mode of action of antagonism, chitinase activity and siderophore production of active isolates were ruled out.
MATERIALS AND METHODS
Bacterial and fungal strains: Isolates of Streptomyces designated as S2 and C were used in this study. The strains were isolated from a soil sample collected from a wheat field at Kerman, Iran as described by Bonjar et al., (2005) and maintained lyophilized before use. Bacteria were revived and cultured on MYA medium containing 10 g Malt extract, 4 g Yeast extract, 4 g Glucose and 15 g agar, adjusted to pH 7.2 and incubated at 29°C for 7-10 days. These cultures were stored at 4°C until used. R. solani AG-4 (Rs1, Rs2 and Rs3 isolates) were provided by Plant Diseases Laboratory of Plant Pests and Diseases Research Institute, Karaj, Iran. Fungal stock cultures were maintained on PDA (Potato Dextrose Agar) at 23°C.
Effect of different temperatures and pH on bacterial growth: The growth behavior of Streptomyces isolates (S2 and C) was evaluated on MYA medium at temperatures of 18, 23, 29 and 37°C and pH ranges of 5.0, 5.6, 7.2 and 8.0.
In vitro antifungal bioassay: Procedure of Yuan and Crawford, (1995) was used to examine the biocontrol activity of Streptomyces isolates S2 and C against R. solani isolates Rs1, Rs2 and Rs3. A loopful of each Streptomyces isolate spores was streaked onto one side of each PDA plate. The plates were incubated at 29°C for 72 h. A 0.45 cm diameter agar plug from the growing margin of fungal colonies was transferred onto the center of the each plate. Fungal plugs were also placed on uninoculated PDA plates separately as uninhibited controls. Cultures were incubated at 23°C. Inhibitory effect of the bacterial isolates on fungal mycelium growth was estimated by subtraction of the fungal growth radius of a control culture (X° in centimeters) from the distance of the growth in the direction of Streptomyces isolates (X in centimeters) after 5 days in dual cultures.
Viability test of treated fungal hyphae: Fungicidal/ fungistatic effect of the antagonists, at the end of 5th day of paired incubation, mycelial plugs from the edge of fungal colony in the vicinity of inhibitory zones were transferred to fresh medium for evaluation of the viability of the mycelia in that area.
Preparation of inoculum for in vivo studies: Dehulled wheat seeds were soaked overnight in tap water, excess of water was removed and the wet seeds were autoclaved twice at 121°C for 20 min, cooled and used as basal medium. For inoculation, pieces of agar with actively growing hyphae of R. solani were placed 2-3 cm below the surface of the medium in 1 L Erlenmeyer flasks. The flasks were shaken every 2 days. After four weeks of incubation at 23°C, the medium was completely covered with the fungal mat. A mixture of the colonized medium, sterilized corn flour and sterilized sand (5, 5 and 90% w/w, respectively) was used as fungal inoculum for in vivo investigations.
Formulation of streptomyces isolates (S2 and C): A spore suspension of each Streptomyces isolates S2 and C was used for in vivo antagonism test. Spores and mycelia from 3 to 6 well grown MYA plates were scraped off of the surfaces with a sterile spatula and added to 20 mL of sterile physiological serum (0.9% NaCl). Spore suspension of each bacterial isolate was added to sterilized sand and mixed carefully, then the formulation was stored at 42°C for 2 h. One g sample from each isolate formulation was used to make serial dilutions in sterile physiological serum and was cultured on MYA plates for measurement of CFU g-1 sand.
Soil treatment and bioassay study: Sterilized soil was infected with inoculum at a rate of 1:10 w/w. In uninoculated pots mixture of 5% w/w sterilized corn flour and 95% w/w sterilized sand was used instead of inoculum. The infected and uninfected soils were placed in 20 cm diameter plastic pots. For plants treated with both bacteria and fungus or plants treated with only bacteria, 80 g (106 CFU g-1 sand) from each formulation was added to inoculated and uninoculated pots. Eighty gram from sand treated with 0.9% sterile physiological serum was added to uninfected soil as control. Pots were prepared in replicates of three, in all treatments and were arranged in a complete randomized design in the greenhouse at 23°C±1 and cycles of 12 h light and 12 h darkness. After 2 days, each pot was planted with 8 monogerm sugar beet seeds and watered every 2 days. After 14 days the numbers of healthy seedlings were recorded. Two months after planting, the plants were harvested and root volume, dry weight of leave, stem and root were measured.
Chitinase activity: Three 0.45 cm diameter agar plug containing mycelium of each Streptomyces isolates (S2 and C) were placed on the surface of 9-cm petri plates containing 0.4% colloidal chitin (Hsu and Lockwood, 1975) and 1.5% agar at pH 7.2 and incubated at 29°C. After 3, 5, 7 and 10 days incubation, clear zones (>5 mm in diameter) around each colony was recorded and used as indicator of chitinolytic activity. There were three replicates and the experiments were repeated.
Siderophore detection: A procedure of Schupp et al. (1988) was used to siderophore detection. In brief, Whatman No. 1 filter paper soaked in indicator solution containing 1% ammonium ferric sulfate (FeNH4(SO4)2.12H2O) in sulfuric acid was put on 4-days single colonies of S2 and C grown up on Des4 medium containing 2% Dextrin, 2% Manitol, 1.2% L-asparagine, 0.025% L-lysine, 0.01% L-methionine, 0.01% L-threonine, 0.5% CaCO3, 0.025% MgSO4, 0.05% K2HPO4, 0.005% ZnSO4 and 2% agar. After 15 min brown or reddish brown halo around colonies were recorded and used to detect siderophore producing Streptomyces isolate.
Statistical analysis: Analysis of variance was performed based on used experimental design. Mean comparison was carried out using Duncan’s multiple range test. MSTATC statistical package was used for the analyses.
RESULTS
Bacterial growth at different pHs and temperatures: Both bacterial isolates grew well at 29°C in pHs of 5.6, 7.2 and 8.0. The isolate C grew in acidic (pH 5.6), neutral (pH 7.2) and alkaline (pH 8.0) media at 23 to 37°C. The isolate S2 grew at 18 to 37°C in pH 5.6 to 8.0 but did not grow in acidic medium at 37°C. When pH decreased to 5.0, the growth of C and S2 isolates was decreased and inhibited, respectively.
In vitro antagonism test: The in vitro antagonism assays showed that both isolates, S2 and C had inhibitory effect on growth of fungal mycelium. The antagonistic effects were appeared after two days of paired bacteria and fungi incubation and advanced during the days after incubation. Comparison of inhibition zones of S2 and C illustrated that isolate C with an inhibition zone more than 2 cm had nearly two fold more inhibitory effect than S2 with an inhibition zone of less than 1 cm. Since Streptomyces have a relatively lower growth rate on agar plates than most of the fungi, preinoculation (72 h) was done to establish bacterial colonies on the agar surface first. Although, inhibition was appeared after two days of simultaneous culture of bacteria and fungi. The viability tests showed that after 5 days of incubation, mycelium of Rhizoctonia isolates (RS1, RS2 and RS3) taken from peripheral areas of the inhibited colonies facing S2 or C, were culturable on fresh agar medium.
Chitinase activity: Clear zone around S2 and C appeared 3 and 5 days after incubation on chitin agar, respectively indicated that both isolates were able to use chitin as a carbon source. Isolate C grew as fast as isolate S2 on chitin agar but did not appear clear zone before 5 days.
Siderophore production: Reaction between siderophore and iron existed in indicator solution ((FeNH4(SO4)2.12H2O)) creates a colored halo around siderophore producing bacteria.
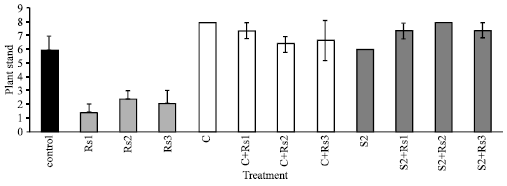 | |
| Fig. 1: | Effect of Streptomyces spp. isolates S2 and C on plant stand of sugar beet seedlings grown in soil infected with R. solani spp. isolate (RS1, RS2 and RS3). Means in each column followed by the same letter are not significantly different from each other according to Duncan,s Multiple Range Test (p<0.05) |
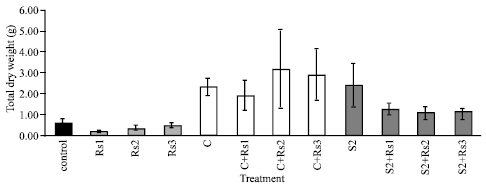 | |
| Fig. 2: | Effect of Streptomyces spp. isolate S2 and C on sugar beet dry matter and root density in soil infected with R. solani spp. isolate RS1, RS2 and RS3. Numbers in each column followed by the same letter are not significantly different from each other according to Duncan,s Multiple Range Test (p<0.05) |
According to that, after covering Des4 medium with Whatman paper soaked in indicator solution the halo appeared around isolate C colonies, but not for isolate S2.
Shelf life of the S2 and C formulation and recovery of bacteria from soil: Density of spores in both formulations of S2 and C was 106 CFU g-1 sand before adding to pots. After 1 months of storage at room temperature (25°C± 2), the density of spores in both formulations have declined to 105 CFU g-1 sand. The level of viability was increased up to 106 CFU/g for S2 and decreased to 104 CFU g-1 for C in samples collected from 3 cm from the soil surface 1 month after treatment with the formulations.
Bioassay study: Soil treatment with S2 and C formulation increased seed germination (16.6 and 25% respectively) from uninfected soil significantly (p<0.05). Streptomyces isolates S2 and C inhibit Rhizoctonia damping-off completely (Fig. 1). All treatments containing bacteria increased shoot and root dry weight and root density compared to control (Fig. 2).
DISCUSSION
Enhancement of fungal biological control agents may improve alternative measures for chemical control management. In this way, several attempts have been made to use the biocontrol approach against major diseases in crop plants.
Several studies have reported the antagonistic effects of Trichoderma spp. (Abada, 1994), Pseudomonas spp. (Shah-Smith and Burns, 1996; Georgakopoulos et al., 2002) and Streptomyces spp (Crawford et al., 1993; Yuan and Crawford, 1995; Paulitz and Belanger, 2001) against Pythium spp. causal agents of sugar beet damping-off. Biocontrol of Rhizoctonia damping off with Streptomyces strains were published for crops, Chinese cabbage by Chung et al. (2005) and tomato by Sabaratnam and Traquair (2002). Also there is a commercial product, Mycostop, driven from Streptomyces griseoviridis has been registered to control of crop damping-off (Lahdenpera et al., 1991).
| Table 1: | Effect of pH (5.0-8.0) and temperature (18-37°C) on growth of Streptomyces spp. isolates S2 and C |
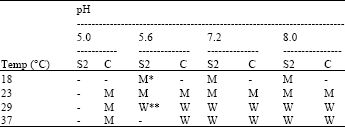 | |
| M*: Moderate growth; W**: Well growth | |
The data presented in this study demonstrated that two Streptomyces isolates (S2 and C) had both in vitro and in vivo antagonistic effects against R. solani, the causal agent of sugar beet damping-off.
Our result showed there is relationship between in vitro antagonism and in vivo disease suppression for potential biocontrol Streptomyces as reported by the others (Yuan and Crawford, 1995; El-Tarabily, 2000).
The in vitro antagonism assays revealed that three pathogenic R. solani isolates were sensitive to the antifungal metabolites released by S2 and C in solid media. Isolate S2 with a smaller zone of inhibition than C was completely able to suppress the disease similar to C based on results from in vivo experiments. It seems that results from in vitro study cannot simply be up scaled to in vivo tests. On the other words, in vitro screening of biocontrol agents should be confirmed using in vivo assays. It may help us to save some beneficial isolates are less effective in in vitro assays.
Soil treatment with both two biocontrol agents increased plant dry matter and root density compared with those were cultured in untreated (with biocontrol agent formulation), infected and uninfected (control) soils (Fig. 2).
Isolates C and S2 showed that differentially promote sugar beet growth. Isolate C increased seed germination (25%) rather than S2 (16%) and total dry matters were more for plants grown is soils treated with C than S2 in infected soil.
Since isolate C, is a siderophore producing and S2 isn't, it may be concluded that a part of efficiency of C is related to siderophore production.
Dicotyledonous plants have been shown to acquire Fe+ from microbial chelates, siderophores (Crowley et al., 1991). Direct utilization of microbial siderophores as iron sources examined and its effect in preventing iron stress and increasing plant biomass reported previously (Wang et al., 1993). On the other hand, siderophore is one of the compounds has been indicated to cause Induced Systemic Resistance (ISR) in plants providing protection against a broad spectrum of phytopathogenic organisms (Bloemberg and Lugtenberg, 2001). Tokala et al. (2002) demonstrated that Streptomyces lydicus WYEC108, an antifungal biocontrol agent, is also a plant growth-promoting bacterium producing hydroxamate-type siderophores. With basis of the results it may point out that siderophore production has a positive correlation with plant growth promotion.
Chitin degradation by isolates S2 and C have been demonstrated on colloidal chitin but its involvement in lyses of living mycelia was not studied. Although, according to De Boer et al. (1998) and Gupta et al. 1995, it seems that the basic mechanism for chitinolytic bacteria against fungi is antibiosis. On the other hand, because mycelial plugs from the fungal colony edge facing S2 and C colonies were recovered on fresh medium, it may be suggested that fungistatic metabolites secretion by antagonist isolates which diffused in solid agar had the most inhibitory effects on R. solani isolates.
Isolates C and S2 grew well at 29°C in pH 7.2, that have been cited as optimal conditions for Streptomyces (Locci, 1994). At 37°C colonies appeared faster and at lower temperature, 23°C both isolates grew moderately. When temperature was decreased to 18°C, the growth of S2 did not change but the growth of C was inhibited (Table 1). Hence both isolates grew moderately to well at temperatures between 25-33°C which R. solani is active (Gallian, 2001) and is suitable for sugar beet. Isolate C at 18°C and isolate S2 in pH 5.0 were inhibited, consequently integration of these two isolates may be more effective to biocontrol of R. solani at some environmental conditions that is not suitable for S2 or C.
Based on these results a biocontrol composite prepared from these two bacterial isolates probably should be viable and active in wide range of temperature (18-37°C) and acidity (pH 5.0-8.0) of planting soils.
Since isolates C and S2 showed inhibitory effects on different genera of fungi (Rhizoctonia, Fusarium and Pyricularia) (unpublished data) seems that these two isolates may be able to suppress all anastomosis groups of Rhizoctonia solani included of the major AG world-wide, R. solani AG-2-2, causes root and crown rot (Scholten et al., 2001) too. Therefore these two isolates have the potential to be developed into commercial biofungicides in control of sugar beet Rhizoctonia diseases.
We hope that further study lead us to identify the mode of action and genes involved in antifungal activity of these isolates.
ACKNOWLEDGMENTS
We thank Reza Ostadsaraie for technical assistance. This work was supported by the Agricultural Biotechnology Research Institute of Iran (ABRII).
REFERENCES
- Abd-Allah, E.F., 2001. Streptomyces plicatus as a model biocontrol agent. Folia Microbiol., 46: 309-314.
CrossRefPubMedDirect Link - Aghighi, S., G.H.S. Bonjar and I. Saadoun, 2004. First report of antifungal properties of a new strain of Streptomyces plicatus (Strain 101) against four Iranian phytopathogenic isolates of Verticillium dahliae, a new horizon in biocontrol agents. Biotechnology, 3: 90-97.
CrossRefDirect Link - Asaka, O. and M. Shoda, 1996. Biocontrol of Rhizoctonia solani damping-off of tomato with Bacillus subtilis RB14. Applied Environ. Microbiol., 62: 4081-4085.
Direct Link - Bloemberg, G.V. and B.J.J. Lugtenberg, 2001. Molecular basis of plant growth promotion and biocontrol by rhizobacteria. Curr. Opin. Plant Biol., 4: 343-350.
CrossRefDirect Link - Bormann, C., D. Baier, I. Horr, C. Raps, J. Berger, G. Jung and H. Schwarz, 1999. Characterization of a novel, antifungal, chitin-binding protein from Streptomyces tendae Tu901 that interferes with growth polarity. J. Bacteriol., 181: 7421-7429.
Direct Link - Chamberlain, K. and D.L. Crawford, 1999. In vitro and in vivo antagonism of pathogenic turfgrass fungi by Streptomyces hygroscopicus strains YCED9 and WYE53. J. Ind. Microbiol. Biotechnol., 23: 641-646.
CrossRefDirect Link - Crawford, D.L., J.M. Lynch, J.M. Whipps and M.A. Ousley, 1993. Isolation and characterization of actinomycete antagonists of a fungal root pathogen. Applied Environ. Microbiol., 59: 3899-3905.
Direct Link - De-Boer, W., P.J.A.K. Gunnewiek, P. Lafeber, J.D. Janse and B.E. Spit et al., 1998. Antifungal properties of chitinolytic dune soil bacteria. Soil Biol. Biochem., 30: 193-203.
CrossRefDirect Link - Georgakopoulos, D.G., P. Fiddaman, C. Leifert and N.E. Malathrakis, 2002. Biological control of cucumber and sugar beet damping-off caused by Pythium ultimum with bacterial and fungal antagonists. J. Applied Microbiol., 92: 1078-1086.
Direct Link - Getha, K. and S. Vikineswary, 2002. Antagonistic effects of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f. sp. cubense race 4: Indirect evidence for the role of antibiosis in the antagonistic process. J. Ind. Microbiol. Biotechnol., 28: 303-310.
CrossRefDirect Link - Gupte, T.E. and S.R. Naik, 1999. Isolation, taxonomic and fermentation studies on a new strain of Streptomyces arenae var ukrainiana producing a tetraene antibiotic. World J. Microbiol. Biotechnol., 15: 545-552.
Direct Link - Jones, C.R. and D.A. Samac, 1996. Biological control of fungi causing alfalfa seedling damping-off with a disease-suppressive strain of Streptomyces. Biol. Control, 7: 196-204.
CrossRefDirect Link - Moussa, T.A.A., 2002. Studies on biological control of sugarbeet pathogen Rhizoctonia solani Kuhn. J. Biol. Sci., 2: 800-804.
CrossRefDirect Link - Nassar, A.H., K.A. El-Tarabily and K. Sivasithamparam, 2003. Growth promotion of bean (Phaseolus vulgaris L.) by a polyamine-producing isolate of Streptomyces griseoluteus. Plant Growth Regul., 40: 97-106.
CrossRefDirect Link - Sabaratnam, S. and J.A. Traquair, 2002. Formulation of a Streptomyces biocontrol agent for the suppression of Rhizoctonia damping-off in tomato transplants. Biol. Control, 23: 245-253.
CrossRef - Schmidt, C.S., F. Agostini, C. Leifert, K. Killham and C.E. Mullins, 2004. Influence of soil temperature and matric potential on sugar beet seedling colonization and suppression of Pythium damping-off by the antagonistic bacteria Pseudomonas fluorescens and Bacillus subtilis. Phytopathology, 94: 351-363.
Direct Link - Bonjar, G.H.S., P.R. Farrokhi, S. Aghighi, L.S. Bonjar and A. Aghelizadeh, 2005. Antifungal characterization of actinomycetes isolated from Kerman, Iran and their future prospects in biological control strategies in greenhouse and field conditions. Plant Pathol. J., 4: 78-84.
CrossRefDirect Link - Tokala, R.K., J.L. Strap, C.M. Jung, D.L. Crawford and M.H. Salove et al., 2002. Novel plant-microbe rhizosphere interaction involving Streptomyces lydicus WYEC108 and the pea plant (Pisum sativum). Applied Environ. Microbiol., 68: 2161-2171.
CrossRefDirect Link - Trejo-Estrada, S.R., I.R. Sepulveda and D.L. Crawford, 1998. In vitro and in vivo antagonism of Streptomyces violaceusniger YCED9 against fungal pathogens of turfgrass. World J. Microbiol. Biotechnol., 14: 865-872.
CrossRefDirect Link - Whipps, J.M., 2001. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot., 52: 487-511.
PubMedDirect Link - Xiao, K., L.L. Kinkel and D.A. Samac, 2002. Biological control of Phytophthora root rots on alfalfa and soybean with Streptomyces. Biol. Control, 23: 285-295.
CrossRefDirect Link - Lahdenpera, M.L., E. Simon and J. Uoti, 1991. Mycostop-A Novel Biofungicide Based on Streptomyces Bacteria. In: Biotic Interactions and Soil-Borne Disease, Volume 23, Beemster, A.B.R., G.J. Bollen, M. Gerlagh, M.A. Ruissen, B. Schippers and A. Tempel (Eds.)., Elsevier, Amsterdam, The Netherlands, pp: 258-263.