
Mustafa Duran
Department of Biology, Faculty of Science and Arts, University of Pamukkale, Denizli-Turkey
Menderes Suicmez
Department of Biology, Faculty of Science and Arts, University of Gaziosmanpaşa, Tokat-Turkey
Murathan Kayim
University of Gaziosmanpaşa, Colloge of Almus, Tokat-Turkey
Cagdas Kaynar
University of Gaziosmanpaşa, Colloge of Almus, Tokat-Turkey
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 5 | Page No.: 848-853







ABSTRACT
Biological characteristics of P. elegans were studied in the coastal water of the Sinop from November 2002 to July 2004. The density low in winter, followed by a general increase as seasonal fluctuation of population density indicated summer and autumn. A cohort of small prawn was identified in May. The largest female measured 69 mm while the corresponding value for males was 60 mm. The proportion of ovigerous adults rose during springs to peak about July the fell to zero by October and ovigerous prawn were taken in six months of the year. A multiple regression of DGR (equation) on temperature and starting length was produced and the growth of juvenile prawn calculated. Females of P. elegans outnumbered males in all months in the study period.
PDF Abstract XML References Citation
How to cite this article
Mustafa Duran, Menderes Suicmez, Murathan Kayim and Cagdas Kaynar, 2006. Preliminary Analysis of the Biological Characteristics of Palaemon elegans (Decapoda, Palaemonidae) in the Coast of Sinop, Black Sea, N. Turkey. Pakistan Journal of Biological Sciences, 9: 848-853.
DOI: 10.3923/pjbs.2006.848.853
URL: https://scialert.net/abstract/?doi=pjbs.2006.848.853
DOI: 10.3923/pjbs.2006.848.853
URL: https://scialert.net/abstract/?doi=pjbs.2006.848.853
INTRODUCTION
Palaemonid prawn Palaemon elegans, Rathke, 1837, is common in European estuaries and elsewhere (Smaldon, 1979). It is essentially a shore species and may also occur in shallow water close offshore. P. elegans is omnivorous, feeding on algae, small crustacean and foraminiferans (Smaldon, 1979). There is very limited data on P. elegans biology in European as light effect on growth and moulting (Dalley, 1980), larval culture (Balestra et al., 1981), mating behaviour (Berg and Sandifer, 1984), temperature effcet on ozmotic and ionik regulation (Ramirez and Taylor, 1985) and a new componenet in macrofauna (Janas et al., 2004) and Turkish waters as larval feeding (Kumlu, 1999) and reproduction biology (Baocinar et al., 2002). This study is focused on growth and population biology of P. elegans in nature, because there are no systematic data on the growth rates of P. elegans of different sizes at a range of temperatures and there is lack of consistent data concerning biological parameters of P. elegans. A number of models of asymptotic growth, Von Bertallanfy, Gompertz and Robertson (Jones, 1976; Cobb and Caddy, 1989), are usually used to describe such non-linear trajectories are often calculated by regression analysis. Linear and quadratic regressions are used in trying to determine the appropriate statistical model for a particular data set.
On this occasion, the present investigation was carried out to study some biological aspect as growth, sex ratio, size frequency, length/weight relationship, maturity and ovigerous prawn of P. elegans inhibiting in the coast of Sinop of Black Sea during 21 months.
MATERIALS AND METHODS
The data were obtained from monthly catches in the coast of Sinop (Black Sea) between November 2002 and July 2004 at the two collecting stations (Fig. 1). The station I was on Akliman (42°00’N, 35°09’E) where, sandy bottom and the stream discharge into the samples area and station II was on Bahceler-Papazlar (42°03’N, 35°02’E) where, sandy-stony bottom with algae. Sampling, at two stations, was made with a bottom trawl net (1.5 m width and 12 mm of mesh pore size), towed for about 30 min at depths of between 2-30 m. Collected specimens were preserved in 70% ethyl alcohol The samples were used to determine species composition, size frequency, length/weight relationship, size at first sexual maturity, daily growth rate and sex ratio of P. elegans.
Total body length was measured from the tip of the rostrum to end of the telson to the nearest mm by rules and separated into length groups and all individuals were weighed to the nearest 0.01 g.
The sex presence (males) or absence (females) of the appendix masculine at the second pair of pleopods was determined.
 | |
| Fig. 1: | Map of the coast of Sinop and stations (St=station) |
Specimens under the minimum sexable size (undetermined) were not considered for this study because of their low number (7.4%).
The regression of log weight on log length gave a linear relationship expressed by log W= log a + log b L. The length/weight relationship were determined separately for the males and females of P. longirostris by the general formula W = aLb, where W is the weight in grams, L the length in cm and a and b are the constant to be calculated.
The growth model is based on field growth rates estimated from size distributions of the monthly samples. At certain times from November 2002 to July 2004 a progression of the mode of the 2 mm length classes from month to month can be clearly followed. In these cases the length frequency data were regrouped into 2 or 1 mm classes and the increment in the modal class was calculated for the class size and over the time period for which the modal shift was distinct. This increase in body was used to calculate a growth rate for that period. Thirty two estimates of growth from the samples date were obtained in this way. In calculating the multiple regressions starting length is taken as the initial modal length (range 17 to 50 mm) and temperature as the mean of the observed sea temperatures for the period (range 6.6 to 22.7°C).
Experimental data were analysed using one-way ANOVA, paired t-test, regression equation and Kruskal-Wallis and any significant difference were determined at a 0.05 probability level using Minitab 13.2 statistical software.
RESULTS
The salinity in the coast of Sinop did not change much throughout the course of this study. Salinity was slightly higher in the summer months (18-20 ‰) than in the winter months (16-18‰). Temperatures ranged from 22.7 to 17.3°C in summer months, from 6.6 to 13.7 in spring months, from 13.9 to 20 in autumn months and from 7.9 to 11.6 in winter months. A significant correlation was found between density, temperature and salinity (One-Way ANOVA (Unstacked), p<0.05).
The pooled mean lengths of P. elegans collected in the samples each months showed consistent decreases in summer in July 2003 and 2004 (Fig. 2). The seasonal pattern of prawn size distribution reveals known summer increase in numbers of cohort small prawn was identified in the samples in June 2003 and in May 2004 (Fig. 3).
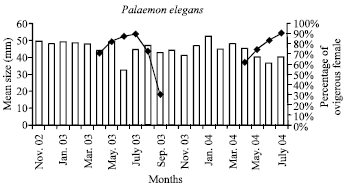 | |
| Fig. 2: | The mean body length (Y1 axis) and percentage of ovigerous female (Y2 axis continuous lines) of P. elegans at the coastal water of the Sinop |
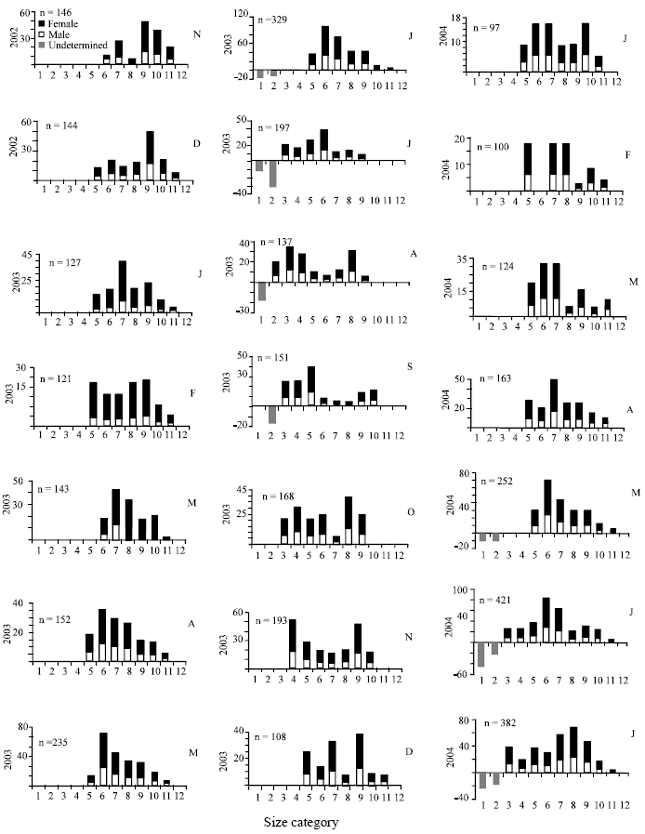 | |
| Fig. 3: | Size frequency distributions of undetermined, females and males of the pooled samples of 21 months for Palaemon elegans. X axis size category where body length (1=13-19, 2=20-24, 3=25-29, 4=30-34, 5=35-39, 6=40-44, 7 = 45-49, 8 = 50-54, 9 = 55-59, 10 = 60-64, 11 = 65-69 mm) |
| Table 1: | Pooled sex ratio of P. elegans for 21 moths from November 2002 to July 2004 |
 | |
Size frequencies between the seasons showed a pronounced seasonal variation each season from November 2002 to July 2004. Size classes showed a regular shift in dominant size class at certain times of the years as the new summer cohort might be seen in May or June to grow in body length through the autumn months to January. Low numbers found in winter, followed by a general increase as seasonal fluctuation of population density indicated summer and autumn (Fig. 3). The highest density 25.7 m-2 in August 2003 and lowest was 0.42 m-2 in January 2004 for station I and the highest density 22 in August 2004 and 0.25 January 2004 for station II was examined throughout the entire study period. A significant different of densities was calculated between stations II and I (Mann-Whitney, p<0.05).
Only a one P. elegans exceeded 69 mm in length. The largest female measured 69 mm while the corresponding value for males was 60 mm and the minimum size at maturity was 28 mm. The largest ovigerous prawn 69 mm and the smallest ovigerous prawn 28 mm were taken in six months of the years. The ovigerous prawn might be carried 470 eggs for 28 mm and 1324 for 69 mm. The proportion of ovigerous adults rose during spring to peak in July both of the years (Fig. 2). In the samples ovigerous prawns were taken in six months of the year. The proportion of ovigerous adults rose during spring to peak about July the fell to zero by November (Fig. 2). There was not a significant difference between station I and II about ovigerous prawns in the samples (Kruskal-Wallis, p>0.05).
Females outnumbered males in most monthly samples in 2003 and 2004. Females predominated all of the year (Table 1). The percentage of female of P elegans 55.2% (1753) and 37.3% (1184) for male and 7.4% (235) for undetermined sex of prawns all over the study period.
The length/weight relationships for each sex W= 0,0030L2.58 for female and W = 0,0030L2.55 for male were calculated.
Daily Growth Rates (DGR) for the field were estimated from the samples length frequency data on 32 occasions, based on the reasonable assumption that the clear progressions of the modal classes during these periods resulted from juvenile growth rate together with mean starting lengths and average temperature for the periods. The field values of DGR regressed on temperature and starting length were significant (for temperature p<0.05, r-sq(adj.) = 73.8%, for starting length p<0.05, r-sq(adj.) = 18.6 %).
A multiple regression of DGR on temperature (T) and starting length (L) was significant DGR = 0.0316 + 0.0076T – 0.00141L p<0.05, r-sq(adj.)= 73.8% (equation).
DISCUSSION
A clear seasonal trend in temperature is obvious as winter/spring lowering temperature. Salinity, temperature and densities intercorrelate. Whether there is much causal interaction between these there variables remains to be seen. Each is tied into a seasonal cycle with the same timing as higher temperatures, higher salinity and higher densities in summer.
A consistent seasonal pattern of abundance in the coast of Sinop has been found over 21 months such as from low number in winter prawn numbers build up during the summer into autumn before dropping into winter as stated by Van den Brink and Van der Velde (1986), Marchand (1981) and Cartaxana (1994, 2003).
Determining a significant difference of densities between stations II and I might be related to the topography of bottom of sample area. The arrival of a new cohort of juveniles from the metamorphosis of the earlier planktonik larval stage is evident in May. This is usually seen in the obvious appearance of an abundance of small (≅ 20 mm) Pelegans in the samples taken at this time. Furthermore, the progressive increase in body size of this cohort through the succeeding summer months as the prawns grow is also clearly discernible. The distinctiveness of the new cohort form May through August implies some degree of synchronicity in the production of larvae, thence juveniles and no substantial further production in summer.
The proportion of berried adult prawn increases form April (around 65% in both years) to August (20%), after this falls to zero by October. The peak of ovigerous females in the coast of Sinop was observed in July. Present data suggests that ovigerous prawn were taken in six months of the year.
In this study, the overall sex ratio was female-biased. According to David and Stuart (2004) deviation from the 1:1 ratio might result from early maturation, higher mortality rates between male and female and some sampling bias with the data collection, possibly due to factors such as the small size of one sex, or relatively habitat selection on invertebrate animals. Van der Brink and Van der Velde (1986) stated that the females P. longirostris found to live longer than the males. It might be results ovigerous females spend prolonged periods for feeding on the bottom or P. elegans female and male use different habitat and they migth be show strong migration and mortality difference between the both sexes.
The length/weight relationships result shows that male and female of P. elegans have fusiform body shape that the confidence for b is less than 3. However condition factors of the males were slightly higher than in the females.
The statistical analysis of the growth rate using linear regression was generally better than that using quadratic regression. This linearity probably relates to the short life span or seasonal migration. In that reason, there appeared no benefit from using an asymptotic Van der Brink and Van der Velde (1986). The particular analysis of these data was to use a simple linear regression. A simple regression of DGR applies positive relationship of temperature and negative relationship of total length. Male growth rate was lower than that of females. However, a slow growth of reproductive female of P. longirostris during the breeding season was observed by Cartaxana (2003) and Hartnoll (1985) also Wickins and Beard (1974) reported that, it is expected that female growth will be reduced during the reproduction period, because a proportion of the available energy is used for the development of oocyts. In fact, average growth rate applicable to field populations of mixed sexes are what is required for most practical purposes. In this reason, average DGR was calculated in this study.
Consequently P. elegans is common in European estuaries but not any data are available on growth of P. elegans. The final discussion will focus particularly on possible life cycle of the P. elegans for these reason, juvenile growth rate is crucial to resolving parts of the life cycle of P. elegans in the coast of Sinop, in particular in determining the duration of the juvenile stage and so how long adults take to develop. The equation is discussed in this work, that estimates of the length of the juvenile stage (from post larvae (≅ 4 mm) to adult, > 40 mm long individuals) were calculated from the equation on the bases of prevailing sea temperatures and prawn body sizes. This was average 11-12 months for P. elegans.
A likely scenario for P. elegans‘s life cycle in the coast of Sinop; first eggs seen on adults around beginning of April will hatch by April or begginning of May and the ensuring larvae develop to juvenile by the middle or end of May. Consequently the first adult will develop form these by April or May. Smaldon (1979) reported that adults may spawn twice times during the year. In general, bigger and older females start egg laying earlier in the year than smaller and young females. Those producing eggs in April will have spawned by April or begginning of May and it is likely that these individuals will produce another clutch of eggs and spawn a second time in that summer.
ACKNOWLEDGMENTS
This study was financed by the Gaziosmanpasa University Research Fund (No. 2002-11). I thank Faculty of Fisheries for supporting the surface temperature and salinity score during the study.
REFERENCES
- Berg, A.B.V. and P.A. Sandifer, 1984. Mating behavior of the grass shrimp Palaemonetes pugio Holthuis (Decapoda, Caridea). J. Crust. Biol., 4: 417-424.
Direct Link - Dalley, R., 1980. Effects of non-circadian light/dark cycles on the growth and moulting of Palaemon elegans reared in the laboratory. Marine Biol., 56: 71-78.
Direct Link - Cartaxana, A., 1994. Distribution and migrations of the prawn Palaemon longirostris in the Mira river estuary (SW Portugal). Estuaries, 13: 685-694.
Direct Link - Cartaxana, A., 2003. Growth of the prawn Palaemon longirostris (Decapoda-Palaemonidae) in Mira river and Estuary, SW Portugal. J. Crus. Biol., 23: 251-257.
Direct Link - David, J.A. and A.W. Stuart, 2004. Sex-ratio evolution in sex changing animals. Evolution, 58: 1019-1027.
Direct Link - Brink, Van den F.W.B. and G. Van der Velde, 1986. Observations on the population dynamics and distribution of the White prawn Palaemon longirostris H. Milne Edwards, 1837 (Crustacea, Decapoda, Natantia) in the Netherlands, with special reference to its occurrence in the major rivers. Archiv. Hydrobiologie, 107: 465-495.
Direct Link