
Abdul Wahab Nassimi
Department of Plant Breeding and Genetics,NWFP Agricultural University, Peshawar, Pakistan
Raziuddin
Department of Plant Breeding and Genetics, NWFP Agricultural University, Peshawar, Pakistan
Naushad Ali
Department of Plant Breeding and Genetics,
Sardar Ali
Department of Plant Breeding and Genetics,
Jehan Bakht
Department of Agronomy, NWFP Agricultural University, Peshawar, Pakistan
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 12 | Page No.: 2333-2337







ABSTRACT
For determination of good combiners in Brassica napus L. genotypes, a study was conducted 8x8 diallel during 2004-05 and 2005-06. F1s and parents were evaluated for combining ability for yield associated traits. Analysis of variance revealed highly significant differences (p≤0.01) for all traits. Components of combining ability analysis exhibited that, GCA was highly significant (p≤0.01) for number of seeds/plant, while significant (p≤0.05) for number of pods/plant and pod length whereas non-significant for 1000 seed weight and seed yield/plant. SCA and RCA effects were highly significant for all traits. GCA effects were higher than SCA and RCA for number of pods/plant and number of seeds/pod indicating that additive gene effects controlled the expression of these traits. The SCA effects were of greater magnitude than GCA effects for 1000 seed weight showing the contribution of non-additive gene effects. RCA effects were greater than GCA and SCA for number of pods/plant and seed yield/plant showing that reciprocal crosses have higher potential than direct crosses for these traits. The parent genotypes NUR1, NUR3, NUR4 and NUR9 and hybrids NUR1xNUR7, NUR2xNUR1, NUR2xNUR8, NUR3xNUR1, NUR3xNUR9, NUR4xNUR7, NUR5xNUR2, NUR5xNUR8, NUR7xNUR4 and NUR8xNUR1 had higher GCA and SCA effects, respectively therefore could be exploited for further selection of high yielding progenies. The overall study revealed the importance of both additive and non-additive genetic variability suggesting the use of integrated breeding strategies which can efficiently utilize the additive as well as non-additive genetic variability.
PDF Abstract XML References Citation
How to cite this article
Abdul Wahab Nassimi, Raziuddin, Naushad Ali, Sardar Ali and Jehan Bakht, 2006. Analysis of Combining Ability in Brassica napus L. Lines for Yield Associated Traits. Pakistan Journal of Biological Sciences, 9: 2333-2337.
DOI: 10.3923/pjbs.2006.2333.2337
URL: https://scialert.net/abstract/?doi=pjbs.2006.2333.2337
DOI: 10.3923/pjbs.2006.2333.2337
URL: https://scialert.net/abstract/?doi=pjbs.2006.2333.2337
INTRODUCTION
Every year, Pakistan imports a huge amount of edible oils for domestic consumption due to lower production of local edible oilseed crops. Non availability of high yielding lines is one of the several factors responsible for this situation. This situation compels plant breeders to develop new high yielding lines of oilseed crops. Rapeseed and mustard is the second major source of edible oil after cottonseed in the country hence enhancement of brassica production can help to reduce the import of edible oils. Advancement in the yield of brassica requires certain information regarding the nature of combining ability of parents available for use in the hybridization program. In addition, information about the nature of gene action involved in expression of quantitative and qualitative traits of economic importance is also required to develop/design desirable lines. According to Zhang (1987) the selection of parents for hybrid cultivar breeding is facilitated by combining ability analysis (combination of GCA and SCA). Information on general and specific combining ability effects is very important in making the next phase of a breeding program. In hybrid breeding, GCA is usually used for parental selection and SCA is used for selection of cross combinations. Therefore, both GCAs and SCAs of female and male parents should be high for hybrid breeding with high yield and high heterosis.
Many workers have reported GCA and SCA effects for yield and yield components in different crops (Muraya et al., 2006; Jan et al., 2005; Attia et al., 2001; Sameena et al., 2000; Khan et al., 1999a, b) whereas critical studies on gene action on yield and yield components in brassica are very few. However, studies on combining ability in relation to combining ability in other traits have been reported by Abercrombie et al. (2005), Satwinder et al. (2000), Kumar et al. (1997), Thakur and Bhateria (1993) and Diwakar and Singh (1993). The main objective of the present study was to identify the best combiners and their crosses on the basis of their general, specific and reciprocal combining ability for yield and its component traits.
MATERIALS AND METHODS
The study was conducted at the experimental farm, Department of Plant Breeding and Genetics, NWFP Agricultural University, Peshawar. Eight Brassica napus L. genotypes viz., NUR1, 2, 3, 4, 5, 7, 8 and NUR9 (NUR for National Uniform Rapeseed Yield Trail) were crossed manually in all possible combinations in a 8x8 diallel fashion during the Rabi season 2004-05. In 2005-06, parents and their 56 F1 hybrids were grown in a randomized complete block design with three replications. All the F1 hybrids and their parent lines were randomly assigned to experimental plots. Each plot comprised four rows of 4 m each with 1 m space between rows and 15 cm between plants.
Data was recorded on number of pods/plant, pod length (cm), number of seeds/pod, 1000 seed weight (g) and seed yield/plant (g) after manual harvesting and threshing. The data were subjected to analysis of variance according to Steel and Torrie (1980). Combining ability effects were computed according to Griffing (1956).
RESULTS
Combining ability is the important tool for the evaluation of line (s) potential performance to be combined with another line (s). Analysis of variance presented in Table 1 showed that for genotypes highly significant differences (p≤0.01) were obtained for all traits studied viz., number of pods/plant, pod length, number of seeds/pod, 1000 seed weight and seed yield/plant. Since genotypic variations were significant for all the characters therefore these variations were partitioned into variation due to General Combining Ability (GCA), Specific Combining Ability (SCA) and Reciprocal Combining Ability (RCA) effects (Table 2).
Number of pods/plant: The mean squares presented in Table 2 indicated that effects due to GCA were significant (p≤0.05) while SCA and RCA were highly significant (p≤0.01) for number of pods/plant (Table 1). According to Table 3, out of 8 parents, 5 showed positive GCA effects for number pods/plant, where the maximum GCA was recorded in NUR1 (20.17), while negative GCA effects were maximum in NUR4 (-23.82). Out of 28 crosses, 16 showed positive SCA effects, the best specific performance with maximum positive SCA effect (54.51) was presented by the cross NUR5xNUR8, while negative SCA effects were the maximum (-67.92) in cross NUR3xNUR9. Data regarding reciprocal effects showed that 17 crosses possessed positive RCA where the maximum value (98.00) was noted in NUR8xNUR1 (Table 3).
Pod length: The mean squares presented in Table 2 shows that effect due to GCA was significant (p≤0.05) while SCA and RCA were highly significant (p≤0.01) for pod length. Pod length is considered one of the major components of seed yield in brassica therefore, positive GCA and SCA were desirable. According to Table 4, out of 8 parents, 2 showed positive GCA effects for pod length. The maximum GCA (111.29) was attained by NUR9, while negative GCA effects were the maximum in NUR2 (-22.69). Data regarding SCA effects showed that out of 28 crosses, 16 showed positive effects, where the best specific performance with maximum positive SCA effect (197.41) was presented by the cross NUR3xNUR9. The maximum negative SCA effects (-112.08) were recorded in NUR5xNUR9. Out of 28 crosses, 16 crosses exhibited positive RCA effects with the maximum positive effects (1.51) being observed in NUR2xNUR1 (Table 4).
Number of seeds/pod: The mean squares showed that effects due to GCA, SCA and RCA were highly significant (p≤0.01) for number of seeds/pod (Table 2). Number of seeds/pod is the most important component of seed yield in brassica therefore; positive combining ability is preferred for it. Data concerning GCA effects (Table 4) pointed that out of 8 parents only 2 showed positive GCA effects for number of seeds/pod. The maximum GCA effects (1.60) were presented by NUR3, while maximum negative effects (-2.20) were in NUR8 (Table 5).
| Table 1: | Mean squares for yield associated traits in Brassica napus L. |
 | |
| ns = non-significant, ** = Highly significant at p≤0.01 | |
| Table 2: | Analysis of variance for combining ability of eight parents and their all possible F1s in Brassica napus |
 | |
| ns = non-significant, * = Significant at p≤0.05 ** = Highly significant at p≤0.01 | |
| Table 3: | General combining ability effects (diagonal values), specific combining ability effects (above diagonal) and reciprocal effects (below diagonal) for number of pods/plant in Brassica napus |
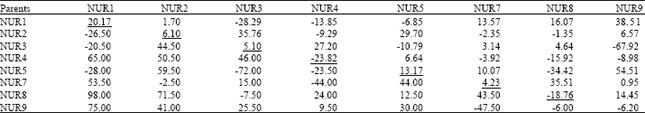 | |
SE of parental value = 4.37, SE for F1 values = 31.24, F-values (EP/EF1) = 7.15, Variance of GCA = 304.684, Variance of SCA = 683.349 | |
| Table 4: | General combining ability effects (diagonal values), specific combining ability effects (above diagonal) and reciprocal effects (below diagonal) for pod length in Brassica napus genotypes |
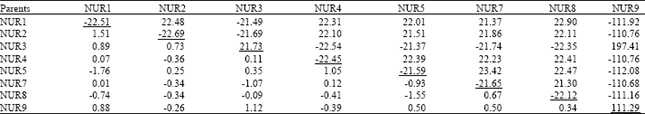 | |
SE of parental value = 0.01, SE for F1 values = 0.01, F-values (EP/EF1) = 1.78, Variance of GCA = 0.168, Variance of SCA = 0.349 | |
| Table 5: | General combining ability effects (diagonal values), specific combining ability effects (above diagonal) and reciprocal effects (below diagonal) for number of seeds/pod in Brassica napus genotypes |
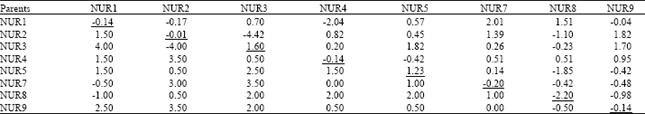 | |
| Variance of GCA = 1.429, Variance of SCA = 2.13 | |
| Table 6: | General combining ability effects (diagonal values), specific combining ability effects (above diagonal) and reciprocal effects (below diagonal) for 1000 seed weight in Brassica napus genotypes |
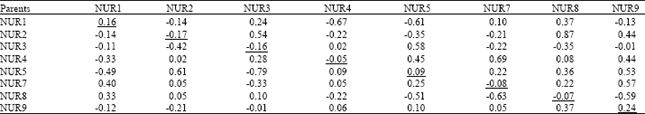 | |
SE of parental value = 0.29, SE for F1 values = 0.36, F-values (EP/EF1) = 1.25, Variance of GCA = 0.027, Variance of SCA = 0.221 | |
Out of 28 crosses, 16 showed positive SCA effects, where the maximum effects (2.01) were presented by the cross NUR1xNUR7, while the maximum negative SCA effects (-4.42) were recorded in cross NUR2xNUR3. RCA effects for number of seeds/plant showed that out of 28 crosses, 24 crosses attained positive effects where the maximum positive effects (4.00) were recorded in NUR3xNUR1.
1000 seed weight: Thousand seed weight is considered as one of the major yield contributing components in brassica. Positive effects for GCA, SCA and RCA were hence desirable. Table 2 showed that mean square values for GCA were non-significant while SCA and RCA were highly significantly different (p≤0.01) for 1000 seed weight. Table 6 showed that, out of 8 parents, only 3 showed positive GCA effects for 1000 seed weight, where the maximum (0.24) being observed in NUR9. While, negative GCA effects were maximum (-0.17) in NUR2. SCA effects indicated that, out of 28 crosses, 17 crosses showed positive SCA effects, where the best specific performance with the maximum positive SCA effects (0.87) was presented by the cross NUR2xNUR8, while the maximum negative SCA effects (-0.67) were found in cross NUR1xNUR4. Positive reciprocal effects were noted in 15 crosses where the maximum positive RCA (0.61) was recorded in NUR5xNUR2.
| Table 7: | General combining ability effects (diagonal values), specific combining ability effects (above diagonal) and reciprocal effects (below diagonal) for seed yield/plant in Brassica napus genotypes |
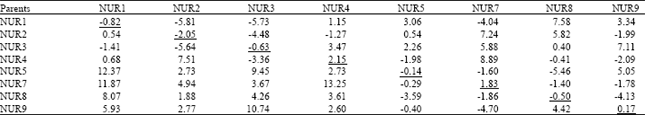 | |
Variance of GCA = 4.385, Variance of SCA = 26.603, SE of parental value = 0.01, SE of F1 value = 0.21, F-value (EF1/EP) = 26.18 | |
Seed yield/plant: Higher seed yield is one of the ultimate objectives of many crops including brassica therefore; positive combining ability effects were desirable. Mean square values for GCA effects were non-significant while SCA and RCA were highly significantly different (p≤0.01) for seed yield/plant (Table 2). Data presented in Table 7 showed that out of 8 parents, 3 showed positive GCA effects for seed yield/plant. The best parent on the basis of the maximum GCA effects (2.15) was found NUR4. SCA effects presented that out of 28 crosses, 14 crosses showed positive effects, where the best specific performance was displayed by the cross NUR4xNUR7 with maximum positive SCA effects (8.89). However, negative SCA effects were greatest in cross NUR1xNUR2 (-5.81). RCA effects were positive in 20 crosses where the highest positive reciprocal effects (13.25) were recorded in cross NUR7xNUR4.
DISCUSSION
Since higher yield in brassica is the major goal of majority of the breeding programs hence improvement in the yield and its components is prerequisite for it. Therefore, positive combining ability is considered desirable for yield associated traits. In our experiment positive GCA, SCA and RCA effects were recorded for all yield related parameters in Brassica napus L. Present results are supported by the earlier findings of Fray et al. (1997) who reported that mean squares due to general and specific combining ability were significant for yield and its components and for oil content. In our study components of combining ability showed that GCA was significant for number of pods/plant and pod length, highly significant for number of seeds/plant whereas, non significant for 1000 seed weight and seed yield/plant. SCA and RCA effects were highly significant for all traits. GCA effects were higher for pod length and number of seeds/pod indicating that these traits were controlled by additive gene action. SCA effects were higher for 1000 seed weight suggesting that this trait was controlled by non-additive gene action whereas; RCA effects were higher for number of pods/plant and seed yield/plant hinting towards the role of maternal effects. From the present results it could be suggested that in Brassica napus L. yield traits were controlled by both additive and non-additive genetic effects. Shen et al. (2005) reported significant variances both for general and specific combining ability for the yield traits. Similarly, Hu et al. (1996) reported that GCA of F1 hybrids was significant for pod length, number of seeds/pod, seed yield/plant and 1000 seed weight. Whereas, SCA was significant for pod length and number of seeds/pod. Similarly, Qian et al. (2003) reported significant general combining ability (GCA) effects for seed yield of hybrids. Davik (1997) stated that GCA of inbred lines and cultivars was significant for pod length, seeds/pod and 1000 seed weight. Our results are similar with the earlier findings of Teklewold and Becker (2005), who found significant Specific Combining Ability (SCA) effects for pods/plant, pod length, seeds/pod, 1000 seed weight and oil content. Krzymanski et al. (1997) revealed significant GCA or SCA effects for seed yield, oil content and some flowering traits. Diwakar and Singh (1993), Thakur and Bhateria (1993) and Kumar et al. (1997) reported significant GCA and SCA effects for yield and component characters indicating that both additive and non-additive gene action were important in the inheritance of these traits. Zhai et al. (2002) reported that in brassica yield/plant and seed oil content were affected by additive gene effects and combination of both additive and non-additive gene effects, respectively. However, Labana et al. (1978) and Dhillon et al. (1990) reported that yield components showed a preponderance of GCA variances compared to SCA variances.
REFERENCES
- Abercrombie, J.M., M.W. Farnham and J.W. Rushing, 2005. Genetic combining ability of glucoraphanin level and other horticultural traits of broccoli. Euphytica, 143: 145-151.
Direct Link - Attia, K.A.E.H., X.Q. Zhong and A.O. Bastawisi, 2001. Combining ability and standard heterosis analysis of two-line system hybrid rice. Pak. J. Biol. Sci., 4: 346-350.
CrossRefDirect Link - Khan, N.I., M. Akbar, N. Iqbal and S. Rasul, 1999. Combining ability analysis in linseed (Linum usitatissimum L.). Pak. J. Biol. Sci., 2: 1405-1407.
CrossRefDirect Link - Jan, M., Farhatullah, Raziudin and H. Ghulam, 2005. Combining ability analysis in sunflower (Helianthus annuus L.). Pak. J. Biol. Sci., 8: 710-713.
CrossRefDirect Link - Khan, J., Shafiullah and Baitullah, 1999. Estimation of combining ability for grain yield and its components in 4 x 4 diallel cross of Maize (Zea mays L.). Pak. J. Biol. Sci., 2: 1423-1426.
CrossRefDirect Link - Muraya, M.M., C.M. Ndirangu and E.O. Omolo, 2006. Heterosis and combining ability in diallel crosses involving maize (Zea mays) S1 lines. Aust. J. Exp. Agric., 46: 387-394.
Direct Link - Qian, W., R. Liu and J. Meng, 2003. Genetic effects on biomass yield in interspecific hybrids between B. napus and B. rapa. Euphytica, 134: 9-15.
Direct Link - Teklewold, A. and H.C. Becker, 2005. Heterosis and combining ability in a diallel cross of Ethiopian mustard inbred lines. Crop Sci., 45: 2629-2635.
Direct Link