
M. Jahanshahi
Department of Anatomy, Gargan University of Medical Sciences, Gargan, Iran
Y. Sadeghi
Cellular and Molecular Research Center, Shahid Beheshti University of Medical Sciences, Tehran, Iran
A. Hosseini
Cellular and Molecular Research Center, Shahid Beheshti University of Medical Sciences, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 8 | Page No.: 1595-1597







ABSTRACT
The hippocampal formation is present in all mammalian species and it is consist of the subiculum, the hippocampus and the dentate gyrus. Apart from principal neurons, the hippocampal formation contains glial cells and various types of interneurons. Glial cells in hippocampus contains the Astrocytes, microglia and oligodantrocytes. Astrocytes play a more active role in neuronal activity, including regulating ion flux currents, energy production, neurotransmitter release and synaptogenesis. Astrocytes are the only cells in the brain that contain the energy molecule glycogen. In this study 5 male albino wistar rats were used. After histological processing. The slides of the brains were stained with PTAH staining for showing the astrocytes. We showed differences in number of astrocytes in different compartment of hippocampus. There is significant difference between CA1 and CA3 and also between CA2 and CA3 areas. We concluded that functional differences can due to structural differences.
PDF Abstract XML References Citation
How to cite this article
M. Jahanshahi, Y. Sadeghi and A. Hosseini, 2006. Estimation of Astrocyte Number in Different Subfield of Rat Hippocampus. Pakistan Journal of Biological Sciences, 9: 1595-1597.
DOI: 10.3923/pjbs.2006.1595.1597
URL: https://scialert.net/abstract/?doi=pjbs.2006.1595.1597
DOI: 10.3923/pjbs.2006.1595.1597
URL: https://scialert.net/abstract/?doi=pjbs.2006.1595.1597
INTRODUCTION
The hippocampal formation is present in all mammalian species. In mammals at developed a three dimensional convoluted structure rather the amphibia in which it has single cortical plate. The volume of hippocampal formation relative to the volume of the whole brain decreases in a phylogenetic sense. However, the relative decrease of hippocampal formation is not caused by a reduction in its size, but by the enormous increase in the size of the neocortex (Rosene and van Hoesen, 1987).
The hippocampal formation consist of the subiculum, the hippocampus and the dentate gyrus (Knowles, 1992 ). These three components have one cell layer encapsulated between two polymorph layers, which is the general organization of the so-called Allocortex (Williams et al., 1995).
The hippocampus can be subdivided into three subfields : The CA1, CA2 and CA3 areas (Amaral and Witter, 1995). The principal cell type in the subiculum and hippocampus is pyramidal neurons. The main cell type in the dentate gyrus is the granule cell. Apart from principal neurons, the hippocampal formation contains glial cells and various types of interneurons, which are situated throughout the hippocampal formation (Williams et al., 1995).
Glial cells in the hippocampus contains the Astrocytes, Microglia and Oligodandrocytes (Williams et al., 1995). In this study, because of the important role of astrocytes in hippocampus, we focused it’s number in different area of hippocampus.
Astrocytes, strategically positioned between the capillaries and neurons, are thought to play a role in neuronal energy metabolism (Pellerin and Magistretti, 2003; Forsyth et al., 1996). Glycogen is localized in the brain almost exclusively in astrocytes (Gruetter, 2003; Tsacopolos, 1996).
Astrocytes and microglia play critical roles in CNS response to and recovery from injury (Rabcheusky, 2002; Bechmann, 1997; Teter, 2002). Astrocytes have been shown to play important roles in nutrient supply, waste removal and axonal guidance. more recent work reveals that astrocytes play a more active role in neuronal activity, including regulating ion flux current, energy production, neurotransmitter release and synaptogenesis. The latter includes the activity of glial cell apposition to synapses and the regulation of synapse elimination by ensheatment [known as glia swelling] (Laming et al., 2000; Teter, 2002).
Astrocytes are the only cells in the brain that contain the energy storage molecule glycogen (Cataldo, 1986). They also contain distinctive 9-nm intermediate filaments composed of a unique protein called glial fibrillary acidic protein [GFAP] (Gimenez, 2000).
MATERIALS AND METHODS
Subjects: Male albino wistar rats (200-250 g) obtained from pasteur institute of Iran were used. Rats were housed in large plastic cage, food and water were available.
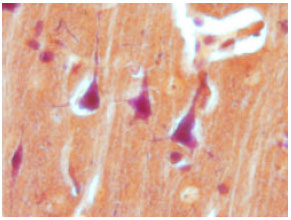 | |
| Fig. 1: | Astrocyte with PTAH staining x 100 |
Animals were maintained under standard conditions and 12 /12 h light/dark cycle with lights on at 7.00 a.m.
Histology: After two weeks, animals were decapitated after ether anesthesia and the brains were removed for histological verification, at first the brains fixed in 10% formalin and two week later, we processed them for embeding with parafine. after embeding, we prepared serial section with 7 μm thickness for each slide. for staining of astrocytes, we used PTAH staining because it is the special staining for astrocyte cells and their processes (Fig. 1).
In this method the astrocytes become blue and the neurons become pink (Fig. 1). Morphometric measurement were carried out using on olympus DP 12 digital camera and BX 51 microscope, selecting a field within the specified cell layer and counting all of the astrocytes shown on the monitor.
Statistical analysis: Data was expressed as mean±SD differences among areas were statistically evaluated using the one-way analysis of variance (ANOVA) probabilities of < 5% (p<0.05) were considered significant.
RESULTS
The mean and SD of the number of Astrocytes in different area of hippocampus (per 75000 μ m2) were 49±17.29 for CA1, 48.82±25.21 for CA2 and 41.95±11.22 for CA3, respectively (Table 1).
There is no significant difference between the number of astrocytes in CA1 and CA2, but there is significant differences between CA1 to CA3 and CA2 to CA3 areas.
We divided hippocampus to three third: Anterior one-third, middle one-third and posterior one-third due to the differ function between anterior and posterior hippocampus (Moser and Moser, 1998).
| Table 1: | Total number of astrocytes in different area of hippocampus |
 | |
| Table 2: | The astrocytes number in different (anterior, middle, posterior) parts of hippocampus |
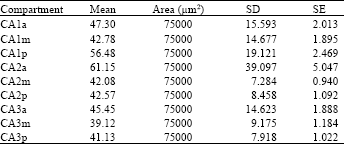 | |
Then we analysed differences between these parts. The mean and SD of astrocytes in different areas (CA1, CA2, CA3) and in different parts (ant., mid., post. ) depicted on Table 2.
There is significant differences between anterior, middle and posterior parts of CA1, respectively and also there is significant differences between anterior and middle parts, anterior and posterior parts of CA2. However no significant difference were observed between middle and posterior parts of CA2.
Also, there is significant differences between anterior and middle parts, anterior and posterior parts of CA3. however no significant difference were observed between middle and posterior parts of CA3.
DISCUSSION
In this study no differences in astrocytes number between CA1 and CA2 areas. but there is significant differences between CA1 and CA2 areas and also between CA2 and CA3 areas, the difference is significant.
Present results resembled to conejo, who worked on the number of astrocytes in rat hippocampus during aging. he showed a significant differences in astrocytes number between CA1 and CA3 area (Conejo, 2003).
We divided hippocampus to three third : Anterior one-third, middle one-third and posterior one-third due to the differ function between anterior and posterior hippocampus (Moser and Moser, 1998 ).
In this study the differences between anterior and middle parts and also between anterior and posterior parts of hippocampus was significant, where, the difference between middle and posterior parts was not significant. This is resemble to Moser and Moser, (1998) and Greicius et al. (2003). They showed different function between anterior one-third and posterior two-third of hippocampus.
Also Pothvizen (2004) showed that the difference function between dorsal and ventral of rat hippocampus with lesion of these areas.
We concluded that functional differences can due to structural differences.
REFERENCES
- Bechmann, I. and R. Nitsch, 1997. Astrocytes and microglia cells in corporate degenerating fibers following enthorhinal lesion. Glia, 20: 145-154.
Direct Link - Cataldo, A.M. and R.D. Broadwell, 1986. Cytochemical identification of cerebral glycogen and glucose-6-phosphatase activity under normal and experimental conditions. II. choroids plexus and ependymal epithelia, endothelia and pericytes. J. Neurocytol., 15: 511-524.
PubMed - Forsyth, R., A. Fray, M. Boutelle, M. Fillenz, C. Middleditch and A. Burchell, 1996. A role for astrocytes in glucose delivery to neurons? Dev. Neurosci., 18: 360-370.
Direct Link - Greicius, M.D., B. Krasnow, J.M. Boyett-Anderson, S. Eliez, A.F. Schatzberg, A.L. Reiss and V. Menon, 2003. Regional analysis of hippocampal activiation during memory encoding and retrieval: FMRI study. Hippocampus, 13: 164-174.
Direct Link - Gruetter, R., 2003. Glycogen. The forgotten cerebral energy store. J. Neurosci. Res., 74: 179-183.
Direct Link - Knowles, W.D., 1992. Normal anatomy and neurophysiology of hippocampal formation. J. Clin. Neurophysiol., 9: 252-263.
Direct Link - Laming, P.R., 2000. Neuronal-glial interactions and behaviour. Neurosci. Biobehav. Rev., 24: 295-340.
Direct Link - Moser, M.B. and E.I. Moser, 1998. Functional differentiation in the hippocampus. Hippocampus, 8: 608-619.
Direct Link - Pellerin, L. and J. Magistrettip, 2003. Food for thought: Challenging the dogmas. J Cereb. Blood Flow Metab., 23: 1282-1286.
Direct Link - Pothvizen, H.H.J., W.N. Zhang, A.L. Jongen-Relo, F. Joram and Y.K. Benjamin, 2004. Dissocition of function between the dorsal and ventral hippocampus in spatial learning abilities of the rat. Eur. J. Neurosci., 19: 705-712.
Direct Link - Teter, B. and J.W. Ashford, 2002. Neuroplasticity in Alzheimer's disease. J. Neurosci. Res., 70: 402-437.
PubMed - Tsacopolos, M. and P.J. Magistretti, 1996. Metabolic coupling between glia and neurons. J. Neurosci., 16: 877-885.
PubMed