
Shokoofeh Hajihashemi
Departments of Biology, Faculty of Science, Azzahra University, Tehran, Iran
Khadijeh Kiarostami
Departments of Biology, Faculty of Science, Azzahra University, Tehran, Iran
Shekoofeh Enteshari
Department of Agriculture University of Valiasr, Rafsanjan, Iran
Azra Saboora
Departments of Biology, Faculty of Science, Azzahra University, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 7 | Page No.: 1370-1374







ABSTRACT
Intra-specific variation in water-soluble carbohydrates (WSC), reducing sugar (RS), photosynthetic pigments and free proline were determined in sixth leaves and roots of two cultivars salt-tolerant (Karchia) and salt-sensitive (Ghods) of wheat (Triticum aestivum) grown in a greenhouse under controlled condition. Plants were irrigated with various levels of salt (0, 75, 150 and 225 mM NaCl) and sprayed with 0, 30, 60 and 90 ppm of paclobutrazol (PBZ) solution. WSC and RS in leaves and RS in roots increased in line with elevating of salinity from 75 to 225 mM of NaCl in both cultivars. A highly significant increase in RS was observed in leaves of cultivar Ghods at level 225 mM NaCl. PBZ treatment increased WSC and RS in leaves and RS in roots in both cultivars, but reduced RS in leaves in cultivar Ghods. Salt stress decreased photosynthesis pigments, significantly in cultivar Ghods, but PBZ treatment increased photosynthesis pigments in plants irrigated with NaCl solution or without. Salt stress and PBZ treatment increased free proline content, but the most significant increase was observed at level 225 mM of NaCl in cultivar Ghods. It may be concluded that PBZ could enhance salt tolerance with increasing osmoprotectants especially in salt-sensitive cultivar.
PDF Abstract XML References Citation
How to cite this article
Shokoofeh Hajihashemi, Khadijeh Kiarostami, Shekoofeh Enteshari and Azra Saboora, 2006. The Effects of Salt Stress and Paclobutrazol on Some Physiological Parameters of Two Salt-tolerant and Salt-sensitive Cultivars of Wheat. Pakistan Journal of Biological Sciences, 9: 1370-1374.
DOI: 10.3923/pjbs.2006.1370.1374
URL: https://scialert.net/abstract/?doi=pjbs.2006.1370.1374
DOI: 10.3923/pjbs.2006.1370.1374
URL: https://scialert.net/abstract/?doi=pjbs.2006.1370.1374
INTRODUCTION
Undoubtedly salinity is a world-wide problem which affecting plant’s growth, yield and its composition. About 25% of world’s total area (including 15% of Iran’s land) is saline. This area contains about 33% of world’s and 50% of Iran’s irrigated lands, respectively (Kamkar et al., 2004). Attempts have been made to overcome the adverse effect of salinity, one of them is application of growth regulators. Paclobutrazol (PBZ) [(2RS, 3RS)-1-(4-chlorophenyl)-4, 4-dimethyl-2-(1, 2, 4-triazol)-pentan-3-ol] is a plant growth regulator and PBZ mediated stress protection is often explained in terms of hormonal changes. Studies showed that PBZ could inhibit gibberllin synthesis, increase cytokinins content, transient rise in ABA and decrease ethylene content (Asare-Boamah and Fletcher, 1986; Fletcher and Hofstra, 1988; Mackay et al., 1990). Some researchers indicated that PBZ reduced the effect of salt stress on guava and grapes (Abou E1 Khashab 1991; Elaidy et al., 1992; Salama et al., 1992). Numerous studies showed that PBZ has a biochemical effect on plants, such as detoxification of active oxygen (Upadhyaya et al., 1990; Kraus and Fletcher, 1994) and increased levels of proline (Mackay et al.,1990), antioxidants (Senaratna et al., 1988) and chlorophyll content (Fletcher and Hofstra, 1988). In spite of the increasing number of studies on PBZ-induced stress protection, a little is known about its effect on the physiological state of salt-stressed wheat plants. Thus we attempt to characterize of the water-soluble carbohydrates (WSC), reducing sugar (RS), photosynthetic pigments and free proline contents under salt stress and PBZ treatment.
MATERIALS AND METHODS
Plant materials and treatment: The experiment was conducted at Azzahra University, Thehran, Iran. Plants were grown in a greenhouse in November 2005, with supplementary light to extend the photoperiod to 16 h per day. Two salt-tolerance (cv. Karchia) and salt-sensitive (cv. Ghods) cultivars of wheat (Pustini, 2001) seeds (provided from the biotechnology and storage of agricultural products center, Karaj, Iran) were sown in 1.5 lit pots containing vermiculite. During the experiment mean temperatures were 24±2°C and 18±2°C, day and night, respectively. The plants were irrigated every 4 days and fertilized every week with the standard Hoagland nutrient solution adjusted to pH 6.0. When two leaves of the plants emerged, they were sprayed with 0, 30, 60 and 90 ppm of PBZ solution. PBZ treatment continued weekly during three consecutive weeks. PBZ treatment, the plants were irrigated with 0, 75, 150 and 225 mM of NaCl solution. Salt stress continued for one month and the plants were leached with tap water after every three irrigations to prevent salt accumulation. Control plants were grown in Hoagland solution and sprayed by water throughout the experiment. All measurements were performed two days after the last day of the salt treatment.
Water soluble carbohydrate (WSC) and reducing sugar (RS): WSC determination was based on the phenol-sulfuric acid method (DuBois et al., 1956). The amount of total RS was measured by Smogyi (1952) method.
Photosynthetic pigments: The photosynthetic pigments were determined by Lichtentaler (1994) method and calculated according to the Lichtentaler and Wellburn (1983) formula.
Proline: Proline was measured in the fresh sixth leaf of plants and measured according to Bates et al. (1973).
Statistical analysis: The experimental design was a random complete block, with three replications. The data were analyzed by the Tukey and Duncan tests’SPSS statistical package to assess significant differences (at p≥5% level) among means.
RESULTS
Carbohydrate: The levels of the WSC and RS in the leaves increased in line with the elevating level of salinity in both cultivars under investigation. But the WSC content in salt irrigated plants was less than control in cultivar Ghods (Fig. 1 and 3). A high significant increase was observed in the RS content in the leaves of cultivar Ghods. The RS content in the leaves was correlated with the salt sensitivity. The PBZ treatment decreased the RS level close to a similar level in control plant in cultivar Ghods (Fig. 3). The PBZ treatment increased the WSC content in the leaves at an elevating level of salinity (Fig. 1). The salt stress decreased the WSC accumulation in the roots of both cultivars. The decrease in the WSC content in cultivar Karchia was highly significant. The PBZ treatment enhanced the WSC content in the roots except at level 30 ppm of PBZ, which showed no considerable difference between levels 75 and 225 mM NaCl solution (Fig. 2). The RS content in the roots of both cultivars grew with increasing salinity. But this increase in all salinity levels was less than in the control plant of cultivar Ghods (Fig. 4). The PBZ treatment without salt stress significantly increased the measured RS in the roots of cultivar Karchia.
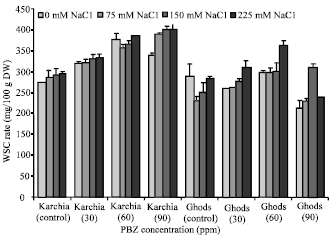 | |
| Fig. 1: | Effects of salt stress and PBZ treatments on the water soluble carbohydrates (WSC) content in the leaves of two cultivars Karchia and Ghods |
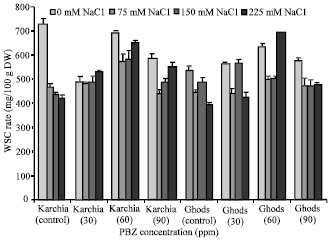 | |
| Fig. 2: | Effects of salt stress and PBZ treatments on the water soluble carbohydrates (WSC) content in the roots of two cultivars Karchia and Ghods |
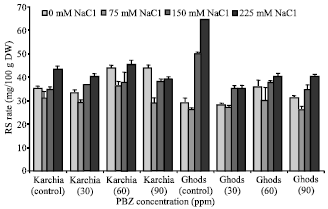 | |
| Fig. 3: | Effects of salt stress and PBZ treatments on the reducing sugar (RS) content in the leaves of two cultivars Karchia and Ghods |
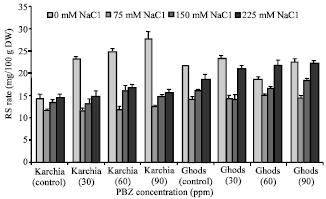 | |
| Fig. 4: | Effects of salt stress and PBZ treatments on the reducing sugar (RS) content in the roots of two cultivars Karchia and Ghods |
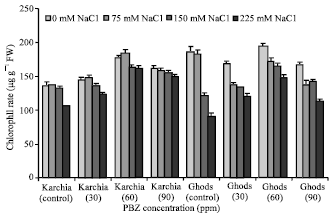 | |
| Fig. 5: | Effects of salt stress and PBZ treatments on the total chlorophyll content in two cultivars Karchia and Ghods |
The WSC level in the roots was greater than in the leaves, but the RS content in the leaves was higher than in the roots. The result suggests the most effective level of PBZ treatment was level 60 ppm of paclobutrazol solution.
Photosynthetic pigments: The total chlorophylls and carotenoids in both cultivars decreased against increasing salinity levels, but this reduction in cultivar Ghods was more significant than in cultivar Karchia (Fig. 5 and 6). The PBZ treatment with or without salt stress enhanced these parameters, significantly in 60 ppm of PBZ treated plants (Fig. 5 and 6). Under salt stress the chlorophyll a/b ratio increased in cultivar Karchia but no significant difference was observed in cultivar Ghods. Results showed that PBZ treatment at level 225 mM NaCl Increased this ratio in both cultivars, with the greatest increase was observed in level of 90 ppm PBZ with a 225 mM NaCl solution.
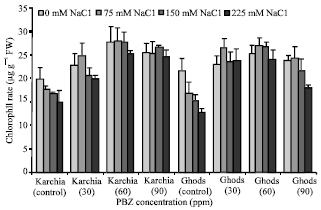 | |
| Fig. 6: | Effects of salt stress and PBZ treatments on the carotenoids content in two cultivars Karchia and Ghods |
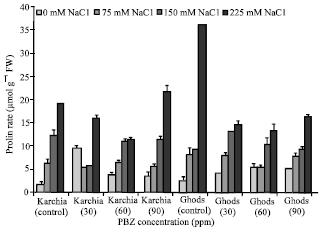 | |
| Fig. 7: | Effects of salt stress and PBZ treatments on the free proline content in the leaves in two cultivars Karchia and Ghods |
Proline: Salt stress and PBZ treatment enhanced free proline. The most significant increase in free proline achieved in cultivar Ghods at level 225 mM of NaCl solution. PBZ treatment had no significant effect in cultivar Karchia, but a significant decrease was observed in cultivar Ghods at level 225 mM of NaCl solution (Fig. 7).
DISCUSSION
The results from this study suggested that under salt stress the WSC content increased in the leaves of cultivar Karchia. But this was less than control plants in the leaves and roots of cultivar and the roots of cultivar Karchia. The PBZ treatment increased this in the leaves and root in both cultivars. Under salt stress, the levels of RS increased in the leaves in both cultivars. This increase was significant in the leaves of cultivar Ghods. The RS content decreased in the leaves of cultivar Ghods at levels near to the control plant. The RS content in the leaves might be used as an indicator in screening wheat genotypes for salinity tolerance. Also it may be the indicat how PBZ treatment increase stress tolerance in plants. High carbohydrate concentration, besides its role in decreasing water potential, contributes both to the prevention of oxidative damage and to the maintenance of the structure of proteins and membrane under moderate dehydration during salt stress. During the course of salt and drought stresses, active solute accumulation of compatible solutes such as amino acids, polyamines and carbohydrates is claimed to be an effective stress tolerance mechanism (Martine et al., 1993; Galiba, 1994; Mckersie and Leshem, 1994; Colmer et al., 1995; Rosa-Ibarra and Maiti, 1995). Carbohydrates also serve as signaling molecules for sugar-responsive genes, leading to different physiological responses, such as defense responses and turgor-driven cell expansion (Koch, 1996; Strum and Tang, 1999). This suggests that increased carbohydrates in PBZ- treated plants increases plant stress tolerance. Photosynthesis, one of the most important metabolic pathways in plants, is a target of salt stress. Decreased the total chlorophyll and carotenoid under salt stress, which in cultivar Ghods was more significant, increased in PBZ- treated plants. It has been reported that several environmental factors, such as drought and low and high temperature can cause an excess amount of toxic oxygen-free radicals (Scandalios, 1993). Carotenoids are known to act as efficient quenchers of triplet chlorophyll (Noguchi et al., 1990) and singlet oxygen (Demmig-Adams, 1990). Elevated amounts of these accessory pigments in PBZ-treated plants should also enhance the capacity to limit the damage caused by species of active oxygen. PBZ enhances the free radical scaveging capacity of treated plants, including the levels of carotenoid, ascorbate and ascorbat peroxidase (Senaratna et al., 1988; Kraus et al., 1995).The highest level of proline in cultivar Ghods at level of 225 mM NaCl may be a suitable indicator for recognizing a salt sensitive cultivar. In addition, PBZ reduced proline content to a level close to cultivar Karchia. Proline acts as an osmoprote- ctant and reduces salt stress damage. It is suggested that the protection and increasing salt tolerance caused by PBZ, especially in salt-sensitive cultivar, was due to mechanisms similar to that of enhanced free-radical scavenging systems and reduced osmotic stress.
Our results suggest that the level of 60 ppm PBZ, is the most effective level in moderating the harmful effect of salt stress. More studies on proteins and enzymes need to indicate how paclobutrazol can protect wheat plants from harmful effect of salt stress.
ACKNOWLEDGMENTS
We are grateful to Dr. Khosrow Manuchehr Kalantari for correcting the English.
REFERENCES
- Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Colmer, T.D., E. Epstein and J. Dvorak, 1995. Differential solute regulation in leaf blades of various ages in salt sensitive wheat and salt tolerant wheat H hopyrum elongatum (Host) love amphiploid. Plant Physiol., 108: 1715-1724.
Direct Link - Demmig-Adams, B., 1990. Carotenoids and photoprotection in plants: A role for the xanthophyll zeaxanthin. Biochim. Biophys. Acta (BBA)-Bioenerg., 1020: 1-24.
CrossRefDirect Link - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Elaidy, A.A., M.I. Salama, A. El-Sammak and A.M. Abou-El-Khashab, 1992. Salt tolerance of Baladi guava seedlings as affected by cultar and ethrel Tanta J. Agric. Res., 18: 373-381.
Direct Link - Galiba, G., 1994. In vitro adaptation for drought and cold hardiness in wheat. Plant Breed. Rev., 12: 115-162.
Direct Link - Koch, K.E., 1996. Carbohydrate-modulated gene expression in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol., 47: 509-540.
CrossRefPubMedDirect Link - Kraus, T., B. McKersie and R. Fletcher, 1995. Paclobutrazol-induced tolerance of wheat leaves to paraquat may involve increased antioxidant enzyme activity. J. Plant Physiol., 145: 570-576.
Direct Link - Lichtenthaler, H.K. and A.R. Wellburn, 1983. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans., 11: 591-592.
CrossRefDirect Link - Mackay, C., J. Hall, G. Hofstra and R. Fletcher, 1990. Uniconazole-induced changes in abscisic acid, total amino acids and proline in Phaseolus vulgaris. Pesticide Biochem. Physiol., 37: 74-82.
Direct Link - Salama, M.I., A.A. Elaidy, A. El-Sammak and A.M. Abou-El-Khashab, 1992. Leaf pigments and nutrient elements content of Roumi Red grape nursling as affected by salinity and some growth regulators. Tanta J. Agric. Res., 18: 382-391.
Direct Link - Scandalios, J.G., 1993. Oxygen stress and superoxide dismutases. Plant Physiol., 101: 7-12.
CrossRefPubMedDirect Link - Upadhyaya, A., T.D. Davis, M.H. Larsen, R.H. Walser and N. Sankhla, 1990. Uniconazole-induced thermotolerance in soybean seedling root tissue. Physiol. Plant., 79: 78-84.
Direct Link