
Najma Yaqoob Chaudhry
Department of Botany, University of the Punjab, Quaid-i-Azam Campus, Lahore 54590, Pakistan
Saima Rasheed
Department of Botany, University of the Punjab, Quaid-i-Azam Campus, Lahore 54590, Pakistan
Pakistan Journal of Biological Sciences
Year: 2003 | Volume: 6 | Issue: 4 | Page No.: 407-412







ABSTRACT
The morphological features of root and shoot of Pisum sativum L., were examined after 30 days. The plant was treated with growth hormones i.e., IAA and Kinetin and heavy metal i.e., Pb (NO3)2. Hormones and heavy metal were used individually i.e., 50 ppm IAA, 50 ppm Kinetin and 50 ppm Pb and in combinations i.e., 50 ppm IAA + 50 ppm Kinetin, 50 ppm IAA + 50 ppm Pb, 50 ppm Kinetin + 50 ppm Pb and 50 ppm IAA + 50 ppm Kinetin + 50 ppm Pb. With applied IAA inhibition in length was observed which was followed by an increase in the diameter in both root and shoot. Extension growth was observed with Kinetin, however, Pb showed inhibition. The number of lateral roots showed increase with IAA and Kinetin but inhibition was observed with Pb. In the mixed doses, length was inhibited. This inhibition was accompanied by expansion in diameter of root and shoot. Moreover, lateral roots showed increase. In the internal morphology of root, an increase in the number of metaxylem elements as well as in the width of these elements was observed with all treatments except Pb. In shoot, the increase in the width of epidermal cells as well as in the cortical region, the number of metaxylem elements, width of these elements and the width of pith region responded positively with all treatments except Pb. Application of heavy metal i.e., Pb(NO3)2 revealed inhibitory effects however, growth hormones i.e., IAA and Kinetin not only balanced the disturbed growth caused by the heavy metal but enhanced the root and stem growth to a certain extent.
PDF Abstract XML References Citation
How to cite this article
Najma Yaqoob Chaudhry and Saima Rasheed, 2003. Study of the External and Internal Morphology of Pisum
sativum L., with Growth Hormones i.e., Indole-3-acetic Acid and Kinetin
and Heavy Metal i.e., Lead Nitrate. Pakistan Journal of Biological Sciences, 6: 407-412.
DOI: 10.3923/pjbs.2003.407.412
URL: https://scialert.net/abstract/?doi=pjbs.2003.407.412
DOI: 10.3923/pjbs.2003.407.412
URL: https://scialert.net/abstract/?doi=pjbs.2003.407.412
INTRODUCTION
The existence of many different types of hormones provide an array of regulatory controls through which differential growth can be achieved (Taiz and Zeiger, 1991). IAA and Kinetin are naturally occurring growth hormones, which show some difference in their mode of action but at the same time they bring about some morphological and physiological changes (Kato, 1958 ; Saunders, 1991). Auxins usually affect other processes in addition to cell elongation (Mauseth, 1991; Raven, 1992; Salisbury and Ross, 1992; Arteca, 1996). According to Davies (1995) auxins stimulate cell elongation, cell division, initiation of cambium and early differentiation of xylem and phloem. IAA metabolism is an essential factor in the regulation of circadian growth of floral stems of Arabidopsis (Jouve et al., 1999). Tuominen et al. (1997) reported that a radial concentration gradient of IAA is related to secondary xylem development. Cytokinins have the ability to induce cell division in the presence of auxin and also promote bud and root formation (Cleland, 1996). Kinetin ordinarily inhibits elongation of stem (Fox, 1964). Furthermore, cytokinin concentration is higher in meristematic regions and areas of continuous growth such as root, young leaves, developing fruits and seeds (Salisbury and Ross, 1992; Raven, 1992). Kinetin was found to be more effective in the yield of soybean (Fatima and Bano, 1998). Khan (1971) reported that high concentration of Kinetin determined the emergence of shoot. Heavy metals are generally toxic and have some physiological effects on plants. Moreover, they are stored or incorporated in living tissues (Botkin and Keller, 1995). Lead binds to essential enzymes and cellular components and inactivates them (Cunning and Sago, 1995). Among heavy metals, Pb is highly toxic to plant growth and delays the germination (Iqbal et al., 2000).
MATERIALS AND METHODS
The present work deals with the study of effect of growth hormones i.e., IAA and Kinetin and heavy metal i.e., Pb [Pb(NO3) 2] on the external and internal morphology of root and shoot of Pisum sativum L. The concentrations were 50 ppm IAA, 50 ppm Kinetin and 50 ppm Pb. In combinations, the composition used was 50 ppm IAA + 50 ppm Kinetin, 50 ppm IAA + 50 ppm Pb, 50 ppm Kinetin + 50 ppm Pb and 50 ppm IAA + 50 ppm Kinetin + 50 ppm Pb. The seeds were grown in petriplates in growth chamber for 30 days in the month of February. 10 ml of each hormone as well as 10 ml Pb was added to the petriplates. The petriplates were kept in dark till germination, approximately 3 days. After that they were given 16 hours of light and 8 hours of darkness and were also given 10 ml of nutrient solution (Hewitt, 1963) alternatively with water. In external morphology, the parameters studied were: 1) length and diameter of root and shoot, 2) number of lateral roots, 3) fresh and dry weight of root and shoot. In order to study the internal morphology, 1 cm long portions of root and shoot were fixed in Corney=s modified fluid. After the removal of air, the fixed material was first dehydrated in an ascending series of water ethyl alcohol mixture, cleared in tertiary butyl alcohol, infiltrated and embedded in paraffin wax. The embedded material was processed with a rotary microtome in the usual manner and transverse sections were cut (10-15 μm). These sections were stained with safranine and fast green, washed in clove oil and mounted in Canada balsam. In the internal morphology, the cells of cortical, xylem and pith region were studied. Data of treated plants was recorded and compared with control. Emphasis was mainly on the promotion or inhibition of cell division, following the treatments. All observations were subjected to statistical analysis (Steel and Torrie, 1981).
RESULTS
Root: In this study, the root of Pisum sativum L., showed inhibition in length with 50 ppm IAA which being 28% in comparison to control. Contrarily, Kinetin showed positive effect. Applied 50 ppm Pb showed a well marked inhibition which was 39%. The mixed doses showed decrease in length when compared with control. The diameter of root showed decrease in length when compared with control. The diameter of root showed a significant increase with IAA and Kinetin i.e., 56 and 25%, respectively. On the other hand, 50 ppm Pb registered a negative effect. The mixed doses of 50 ppm IAA + 50 ppm Pb and 50 ppm Kinetin + 50 ppm Pb showed an increase over control, however, remarkable increase was observed with 50 ppm IAA+50 ppm Kinetin and 50 ppm IAA+50 ppm Kinetin + 50 ppm Pb, which was 93 and 87%, respectively. The number of rootlets showed an increase with all the treatments except Pb, which showed inhibition which was 33% over control. The fresh and dry weight showed inhibition with all the treatments except Kinetin, which showed an increase (Table 1).
In the internal morphology, the root was tetrarch. This condition remained constant in the treated plants. The number of cortical layers were 16 in control as well as in Pb treated plants, however, in the rest of the treated plants a layer was increased. The number of metaxylem elements showed an increase with IAA and Kinetin as compared to control. Pb, however, showed no change. The mixed doses of IAA+Pb and Kinetin+Pb registered a negligible increase. Furthermore, the doses of IAA+Kinetin and IAA+Kinetin+Pb showed an increase, which was 28 and 21%, respectively. The width of metaxylem elements showed a well marked increase with all treatments except 50 ppm Pb, which registered a negative effect (Table 3).
Stem: The stem showed an inhibition in length with all treatments except Kinetin. The 50 ppm Kinetin showed an increase in length as compared with control. The diameter of stem showed expansion with 50 ppm IAA and 50 ppm Kinetin. The rest of the treatments also showed some increase, however, 50 ppm Pb showed inhibition. The fresh weight of stem registered an inhibition with IAA and Pb. However, the applied 50 ppm Kinetin showed a well marked increase of 15% over control contrarily the mixed doses showed an inhibition in fresh weight. The dry weight of stem showed decrease with all treatments except Kinetin, which showed a negligible increase of 10% over control (Table 2).
In the internal morphology, the width of epidermal cells showed an increase with all treatments as compared to control. However, Pb registered some inhibition. The number of cortical layers remained constant with all the treatments i.e. 14. An increase in the width of cortical region was observed with 50 ppm IAA and 50 ppm Kinetin i.e. 17.03 and 14%, respectively. However, 50 ppm Pb showed an inhibition.
| Table 1: | Effect of growth hormones and heavy metal on external morphology of root |
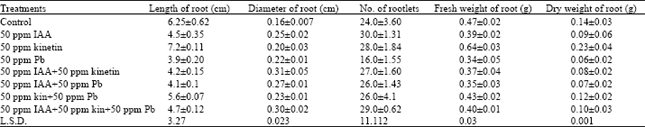 | |
| Table 2: | Effect of growth hormones and heavy metal on external morphology of shoot |
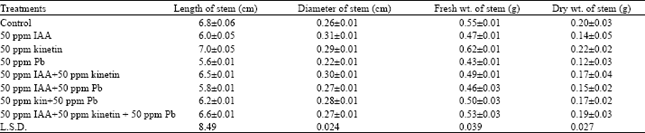 | |
| Table 3: | Effect of growth hormones and heavy metal on internal morphology of root |
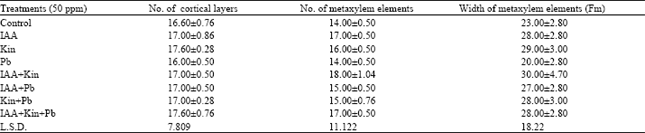 | |
| Table 4: | Effect of growth hormones and heavy metal on internal morphology of stem |
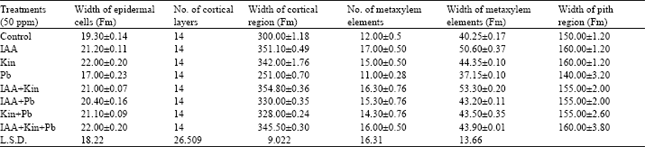 | |
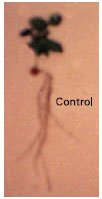 | |
| Fig. 1: | P. sativum L. control |
Likewise the mixed doses also showed an increase in the width of cortical region but the dose of 50 ppm IAA+ 50 ppm Kinetin registered a well marked increase of 18.2% over control (Table 4).
The maximum number of metaxylem elements were observed with 50 ppm IAA, 50 ppm IAA + 50 ppm Kinetin and 50 ppm IAA + 50 ppm Kinetin + 50 ppm Pb as compared with control. The rest of treatments also showed an increase except 50 ppm Pb, which showed inhibition. The width of metaxylem elements showed an increase with 50 ppm IAA i.e. 25.7%.
 | |
| Fig. 2: | Effect of IAA + Kinetin |
Similarly, 50 ppm Kinetin also showed an increase. However, 50 ppm Pb registered negative effect. The mixed doses showed an increase in the width of metaxylem elements but 50 ppm IAA + 50 ppm Kinetin showed well marked increase of 32.42% (Table 4). The width of pith region showed an increase with all treatments except Pb, which registered some inhibition (Table 4).
 | |
| Fig. 3: | Effect of IAA + Pb |
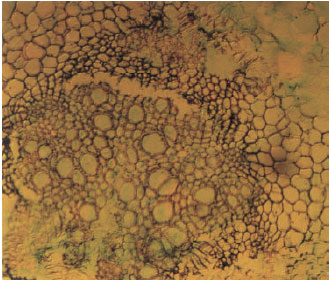 | |
| Fig. 4: | Effect of IAA + Kinetin on xylem elements |
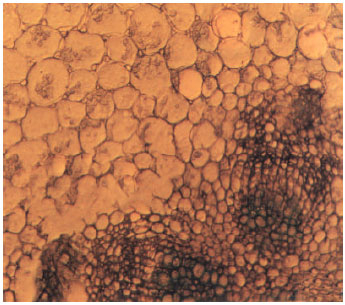 | |
| Fig. 5: | Effect of IAA + lead |
DISCUSSION
Root: In the external morphology, the application of extraneous IAA showed inhibition in root length in comparison to control (Fig. 1). Similar reports are given by number of workers (Votrubova and Vatruba, 1981; Saxena and Singh, 1982; Evans, 1954; Pilet and Saugy, 1985).
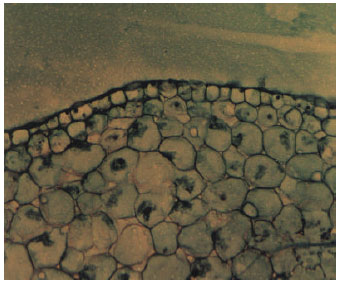 | |
| Fig. 6: | Effect of Kinetin + Pb on cortical region and epidermal cells |
However, the diameter fresh and dry weight and the number of rootlets showed an increase with IAA. The increase in diameter may be due to the radial expansion (Jernstedt, 1984). The increase in the number of rootlets may be attributed to the auxins which enhance initiation of root meristem and consequently increase the number of lateral roots (Audus, 1959). Likewise, applied Kinetin, showed increase in all parameters.
The length of root showed increase with Kinetin. Zadoo (1986), Makarova et al. (1988) have reported that Kinetin promotes growth in all parameters. The application of lead, showed inhibitory effects. This may be due to the toxic effects of lead which is well known for displacing of biological processes (Ivanov et al., 1997). The equal mixed doses showed an inhibition in length which was followed by an increase in diameter, fresh and dry weight and the number of rootlets (Table 1; Fig. 2, 3). These results show that IAA and lead have inhibitory, effects on length, similar results have been reported by Vanderhoef and Key (1968) while working on excised soykean hypocotyls. On the other hand, IAA and Kinetin showed increase in diameter. Katsumi (1962) reported that IAA and Kinetin stimulate expansion growth.
In the internal morphology, an increase in the number of cortical layers was observed i.e. one layer was increased as compared with control. This may be due to enhanced cell division (Cleland, 1996). However, lead showed no effect on cortical layers. This shows that lead inhibits growth parameters (Hassanein and Shehata, 1999). The application of IAA and Kinetin showed an increase in the number of metaxylem elements as well as in their width. Wareing and Roberts (1956) reported wider metaxylem vessels with the application of auxins. Zadoo (1986) and Makarova et al. (1988) reported similar results with Kinetin. Contrarily, lead showed no change in the number of metaxylem elements and inhibition was observed in the width. According to Iqbal et al. (2000) lead inhibits plant growth. The mixed doses, showed an increase in the number and width of metaxylem elements. This may be due to the combined effect of both hormones. As Davies (1995) and Chaudhry and Khan (2000) reported that Kinetin is known to promote expansion alongwith IAA (Table 3; Fig. 4).
Stem: In the external morphology, the application of IAA inhibited length, similar reports have been given by Adams and Ross (1983). IAA inhibits length (Davies, 1995), however, this inhibition is followed by a corresponding increase in diameter (Chaudhry and Rashid, 2000). The fresh and dry weight showed decrease which may be attributed to the decrease in length (Pilet and Saugy, 1985). Applied Kinetin showed some increase in length as well as in diameter. Cytokinins are well known for their action on expansion growth (Makarova et al., 1988). Katsumi (1962) has reported that Kinetin increases fresh and dry weight this report is similar to the present observations. The inhibitory effect of lead was observed in all parameters. These observations were in harmony with the observations of Hassanein and Shehata (1999). In the mixed doses, inhibition in length was observed which may be attributed to the mixed dose of lead (Table 3; Fig. 2, 3). However on the other hand, diameter showed an increase with mixed doses over the control, similar reports were given by Mauseth (1991) and Raven (1992).
In the internal morphology, the number of cortical layers remained same with all the treatments. This shows that no cell division is promoted in this region following treatments. Applied IAA showed an expansion in the width of epidermal cells (Table 4; Fig. 6). Chaudhry (1997) has observed similar effect. The positive effect of Kinetin was observed in all parameters undertaken (Table 4; Fig. 6) which is well known for causing cell expansion (Makarova et al., 1988) and cell enlargement (Powell and Griffith, 1960). An inhibition in all parameters was observed with lead, which is well known for its inhibitory effect (Iqbal et al., 2000). The mixed doses showed positive effect. This may be attributed to the combined effect of IAA and Kinetin (Davies, 1995 and Raven, 1992). However, with mixed doses no regular pattern was observed (Table 4; Fig. 5, 6). In all the parameters observed the antagonistic effect of lead nitrate has been registered.
REFERENCES
- Adams, P.A. and M.A. Ross, 1983. Interaction of indoleacetic acid and gibberellic acid in the short-term growth kinetics of oat stem segments. Plant Physiol., 73: 566-568.
Direct Link - Jernstedt, J.A., 1984. Root contraction in hyacinth. I. Effects of IAA on differential cell expansion. Am. J. Bot., 71: 1080-1089.
Direct Link - Khan, A.A., 1971. Cytokinins: Permissive role in seed germination. Science, 171: 853-859.
CrossRefDirect Link - Wareing, P.F. and D.L. Roberts, 1956. Photoperiodic control of cambial activity in Robimia pseudacacia L. New Phytol., 55: 356-366.
CrossRefDirect Link