
Afsar Mian
Department of Biological Sciences, University of Arid Agriculture, Rawalpindi, Pakistan
Pakistan Journal of Biological Sciences
Year: 2003 | Volume: 6 | Issue: 14 | Page No.: 1282-1295







ABSTRACT
Phytosocio logical analysis carried out in 53 stands, created in habitat of wintering Houbara in Balochistan, suggests presence of at least 90 species, appearing in different combinations in 21 communities distributed under different conditions of soil, temperature and altitudes. The constancy of appearance of most of the species is low (> 20% only for Rhazya stricta, Anabasis sp., Pennisetum dichotomum, Calligonum comosum and Haloxylon ammodendron). There is a good degree of intra-stand homogeneity and most of the stands have < I I species. Shrubs constitute the only regular layer, herbs appearing during some part of the year and no regular tree layer. Vegetation appears in patches. Vegetative cover ranges between 0.87 and 34.17 % but generally it ranges between I and 15%. Most of the species are vigorous, reproducing sexually and/or a sexually and exhibit different degree of xerophytic characters. The phenological cycle exhibited by different species can be explained on temperature and precipitation pattern.
PDF Abstract XML References Citation
How to cite this article
Afsar Mian, 2003. On Biology of Houbara Bustard (Chlamydotis undulata macqueenii) in Balochistan, Pakistan:Phytosociological Analysis of Habitat. Pakistan Journal of Biological Sciences, 6: 1282-1295.
DOI: 10.3923/pjbs.2003.1282.1295
URL: https://scialert.net/abstract/?doi=pjbs.2003.1282.1295
DOI: 10.3923/pjbs.2003.1282.1295
URL: https://scialert.net/abstract/?doi=pjbs.2003.1282.1295
INTRODUCTION
Phytosociological studies in habitat of animal species help in knowing present and future habitat potentials. Such studies are important in placing conservation of the animal species on sounder footings, especially in the species, like Houbara, placing a good degree of reliance on general vegetation for food and shelter. Casual lists of prominent plant species present in different Houbara tracts have appeared in different reports (Cramp and Simmon, 1980; Ali and Ripley, 1983 and Roberts, l991), yet very few studies attempted detailed phytoecological analysis (Goriup, 1983a and Collins, 1983, 1984). Some indirect references to phytohabitat are also available Loughland and Launay, 1994; Launay et al. 1994; Rashid and Nadeem, 1997 and Mian et al.1999.
Present study has been conducted to provide a detailed phytoecological analysis of habitat exploited by Houbara in its wintering grounds in Balochistan, Pakistan. Basic data presented can be used in habitat management programmes and in analysis of influence of different phytoecological factors in controlling Houbara population in different areas.
MATERIALS AND METHODS
Fifty-three stands (large tracts exhibiting visual phytological homogeneity) were established to sample different Houbara tracts of Balochistan (Fig. 1). All the stands were visited during early (October-November), mid (December-Janu -and late (February-March) winter and during different years (1983-84 through 1986-87).
Balochistan (southwestern province of Pakistan, area 343,000 kM2) is traversed by I I principal mountain ranges. The valleys of different sizes are present between these mountain ranges. Houbara exploits broad valleys (some 75,000 kM2 ) during winter (September-March). These valleys can be divided into 5 broad localities, i.e., northern highlands (altitude = 1,0001,700 m), central highlands (6001,200 m), central lowlands (<100 m, eastern parts), southern highlands (600-1,000 m) and southern lowlands (<200 m).
Vegetation was studied by quadrat method following layer-by-layer analysis. Three vegetative layers, i.e., tree (>2 rn tall, quadrat = 40 × 10 m); shrubs, tall grasses and small trees (0.15-2.00 m tall, quadrat =10 × 10 m); herbs and small grasses (<0.15 m tall, quadrat = I × I m), were recognized following Kayani (1984). In each stand, 15-20 quadrats were selected randomly to include all possible microhabitat variation.
Absolute vegetative cover was determined by crown diameter methods (Mueller Dombois and Ellenberg, 1974). Optimal crown diameter of each plant, falling in quadrat area, was recorded separately, and used for calculation of crown area (7r r2; where, n = 3.1429 and r= radius). Absolute cover of each plant species and total absolute vegetative cover in quadrat area was worked out through appropriate pooling of individual crown areas and expressed as percentiles of total area under respective quadrat. General stand cover was calculated as means of quadrat data. Community was named after the species having dominant cover. Plant species were identified/named following Burkill (1909) and Stewart (1972).
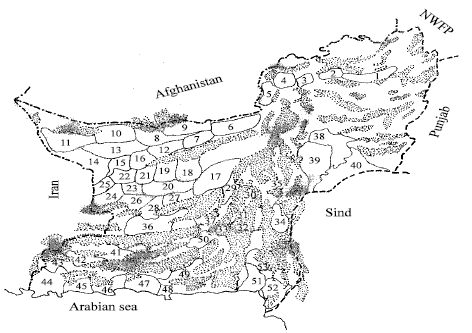 | |
| Fig. 1: | Line sketch of Balochistan (Pakistan) showing the tentative location of the stand areas |
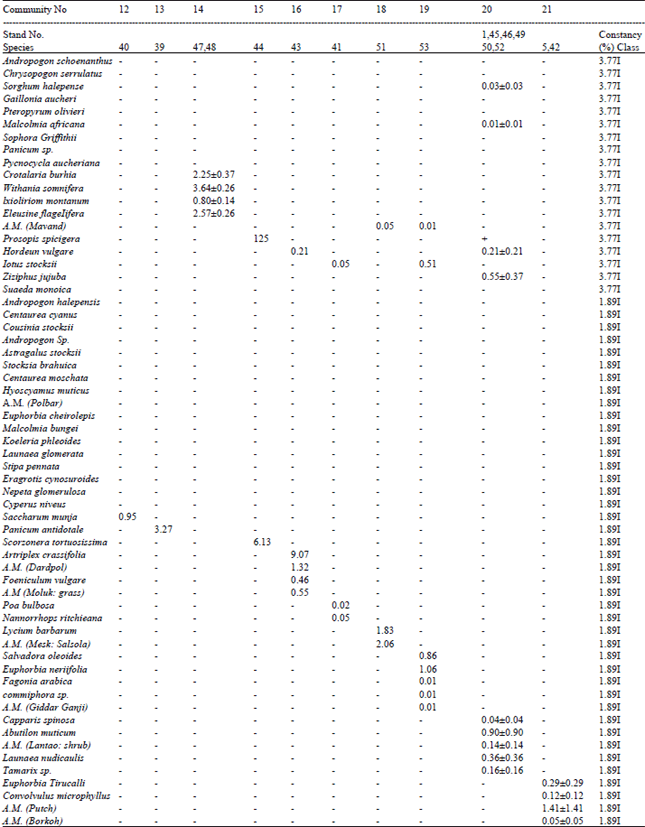
Constancy of appearance of each species was worked out by dividing the number of stands showing the plant species by total number of stands studied and expressed in percentiles. Each species was assigned to one of the five constancy classes (class I= <21% constancy, II= 21-40, III= 41-60, IV= 61-80, V=>80%, Mueller-Dombois and Ellenberg, 1974.
Field notes were maintained on general distribution of prevalent vegetation (physiognomy). Each stand was classified into one of the sociability classes, recognized by Braun-B lanquet (1951) class I = plants scattered as individual stalks, 11 = small groups of plants, III = small scattered patches or cushions, IV = large unbroken cushions, V = unbroken mat of vegetation).
Vigour of each species was recorded, giving due consideration to general vegetative state under different stress conditions and its potentials of completing life cycle. Each species was classified into one of the vigour (vitality) classes following Mueller-Dombois and Ellenberg (1974) class I= well developed, regularly completing life cycle; 11 = vigorously growing plants and mainly spreading through vegetative propagation; III = never completing life cycle and spreading vegetatively; IV = occasionally appearing from seeds, but do not increase in number).
Field records were maintained on vegetative form of each species, which was placed in one of classes (phanerophytes = shrubs, trees and vines with buds located on upright shoots at least 25 cm above surface; chamaephytes = plants with buds located from ground surface to 25 cm above it; hemicryptophytes= plant with perennial shoots and buds located close to surface; geophytes= plants with perennating parts buried in substratum (Mueller-Dombois and Ellenberg, 1974).
Phenological records, such as, degree of foliage, flowers and seeds/fruits, were recorded through a visual impression. Data such collected were synthesized to evolve a general pattern for the broad locality.
The general topography, altitude and broad soil type (sand, silt, clay, depending upon general visual impression) of each stand was recorded in the field. The data on precipitation and temperature were adopted from Champion et al. (1965).
RESULTS
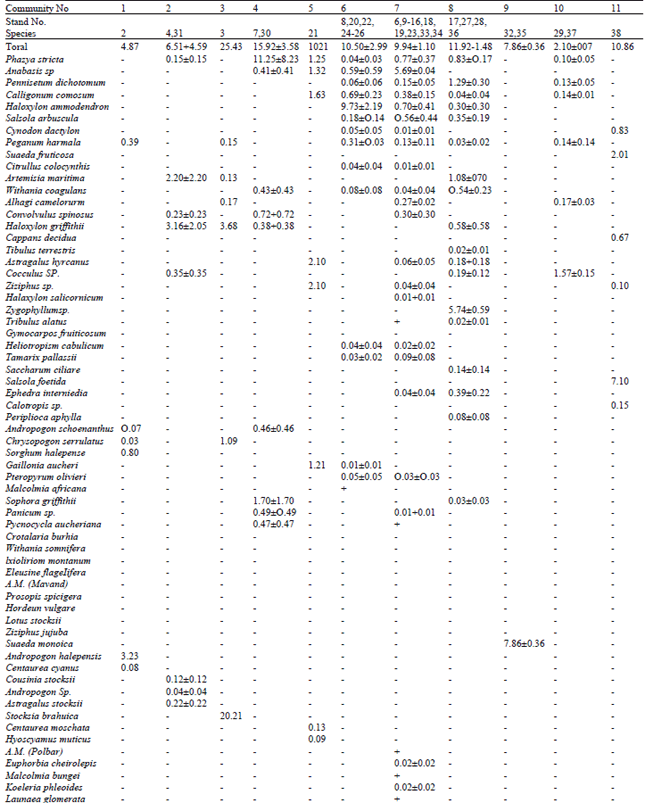
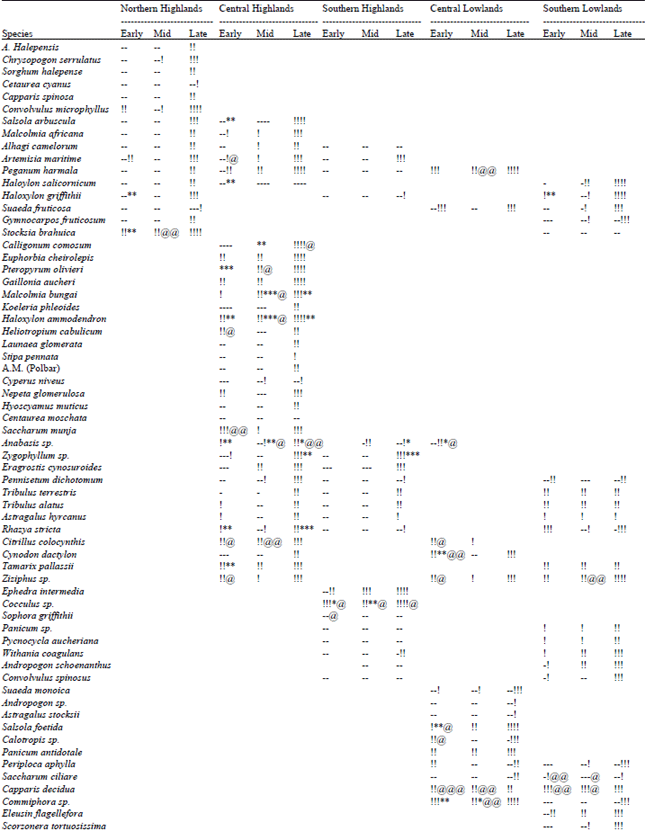
Phytoecological characters: There were at least 90 species of angiosperms (dicot = 74, monocot = 16) in bustard tracts (Table 1). Individual stands had few species' (I-11, 14 in only one stand). Constancy of appearance of different species (Table 1, last column) was generally low. Only Rhazya stricta fell in constancy class 111, while 4 species (Anabasis sp., Pennisetum dichotomum, Calligonum comosum and Haloxylon ammodendron) were in class 11. All other species fell in class 1 (42 represented in one, 17 in two, 8 in three stands). This suggests intra-stand homogeneity and inter-stands heterogeneity.
Vegetative cover was generally low and ranged between 0.87 and 34.17% in different stands. However, more commonly it remained between 5 and 15%.
The sociability indicated that vegetation was mainly present as small groups of plants (class 11, 87% of stands). It also appeared as small cushions of plants (class 111, 12%) or as isolated stalks (Class 1, 1%).
Stratification was poor. Only shrubs constituted a regular layer, herbs and small grasses appearing occasionally and during some part of the year.
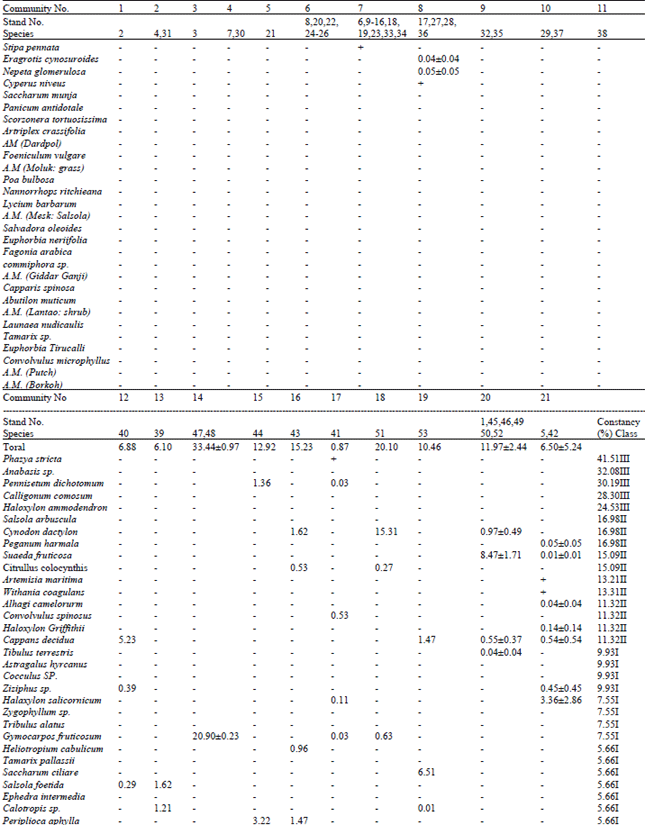
| Table 1: | Distribution of absolute cover (%) amongst different species in different vegetative communities recognized from Houbara wintering tracts in Balochistan community numbers correspond with the text. and stand numbers with Fig. 1 |
   | |
Isolated plants and/or small groups of trees represented tree layer. The major part of the species was phanerophytes (85%), mainly represented by shrubs (68%), though some were with herbaceous stem (I I%), trees (6%) and vines (2%). Chaernophytes constituted some 13% of the species. Hemicryptophytes, cryptophytes and tharophytes bore a very limited appearance during winter. All the species exhibited different degree of xerophytic characters, yet succulents were few.
All the species exhibited variation in their vegetative state during different parts of winter (phenological cycle), as also between years and area (variable temperature, precipitation and soil moisture), yet generally these were vigorous. Under persisting drought (hitting the major parts between 1983 and 1987), however, a good proportion of plants of almost all the perennials started giving dry look during 1986-87, when there was limited foliage. Majority of the species was placed in vitality class 11 (91%, reproducing both through vegetative propagation and seeds), while some 9% were in class I (propagating through seeds only).
Vegetation communities: On the basis of dominant cover of the species, 21 vegetative communities were recognized with following composition (Table 1) and distribution.
Andropogon halepensis: A. halepensis contributed dominant cover (66.3% of community cover) in the foothills of Toba Kakar Range (northern highlands, alt. 1,700 m). Stand mainly represented a flat plain with widely dispersed watercourses and vegetation appearing as scattered stalks along the edges. Soil was silt mixed with sand. Community presented an association of 6 species. Four species of tall grasses (A. halepensis, A. schoenanthus, Chrysopogon serrulatus, Sorghum halepense) shared major cover (90.21%), while two dicots (Peganum harmala, Centaurea cyanus) contributed a minor part.
Haloxylon griffithii: In two isolated stands, H. griffithii provided the dominant cover. Both the stands had silty but partly stabilized soil in mild hilly undulates. In southern reaches of northern highlands (alt. 1,700 m), a tract represented this community with a relatively good vegetative cover. Here the dominant species was associated with Artemisia maritima. Cousinia stocksii contributed a minor part of total cover. In a stand located in the northern reaches of southern highlands (alt. 600 m), Cocculus sp. appeared as a second dominant with H. griffithii. The other 4 species had only a minor contribution in the stand cover. The overall stand cover was low, vegetation mainly appearing along watercourses.
Stocksia brahuica: Undulating highland hilly valley (alt. 1, 700 m) having loose stabilized soil represented this community. Watercourses were deep and with sharp edges, especially towards foothills. Vegetation produced a high absolute cover. S. brahuica was present along edges of water courses, while H. griffithii dominated the ridges. S. brahuica, though bore a dominant community cover, yet it was distributed as scattered large shrub. Other 4 species had a minor contribution in community cover, except Chrysopogon serrulatus.
Rhazya stricta: R. stricta provided dominant cover in two isolated stands sharing stabilized soil in mild hilly slope flat. In central highlands (Chagai, alt. 900 m) it was represented by a pure stand of evenly distributed R. stricta having a high vegetative cover. In northern reaches of southern highlands (alt. 600 m) it suggested an associated of 9 species. R. stricta shared dominant cover with Sophora griffithii. Other 7 species were almost equally distributed in the stand, though Convolvulus spinosus was better represented.
Astragalus hyrcanus: Community was located in hilly undulates of western Kharan (alt. 600 m). Soil was relatively stabilized with a loose texture. Watercourses had sharp and broken edges, especially towards foothills. Community had a good cover of scattered shrubs and represented an association of 8 species. None of the species provided a dominant cover, yet A. hyrcanus, Ziziphus sp. and Calligonum comosum shared a higher cover. Out of other 5 species, Anabasis sp., Rhazjw stricta and Gaillonia aucheri shared almost equal parts, while Hyoscyamus muticus and Centaurea moschata bore a minor part of total stand cover. The vegetation was sparse in general valley, though better vegetation Was present along heavily eroded watercourses.
Haloxylon ammodendron: This community was distributed over a wide tract of central highland valleys (alt. 600-900 m; Chagai and Kharan). It presented an association of 12 species. Cover was mainly contributed by widely scattered large shrub of H. ammodendron. The constancy of appearance of other species was very low. Most of these appeared in an isolated stand, except for Calligonum comosum (4 stands), Rhazya stricta, Peganum harmala, Salsola arbuscula and Tamarix pallasii (2 stands each). This community was present in extreme deserts, with moderate to high undulating sand dunes.
Anabasis sp.: The community was spread over a very wide area of central highlands (alt. 6001,200 m; Chagai and Kharan) and some parts of southern highlands (Khuzdar). It presented an association of 33 species. Anabasis sp. contributed major shares in total cover (around 75%, range 52100%) in 12 stands, while in 2 it shared the dominant cover with Salsola arbuscula and Convolvulus spinosus, respectively. Other species, associated with the dominant species, generally shared a small part of community/stand covers. Rhazya stricta was widely distributed (64% constancy in community) contributing next highest cover. Three species (Calligonum comosum, Pennisetum dichotomum, Haloxylon ammodendron) had significant contribution in total average cover and were widely distributed. All other species contributed a minor part in total cover and appeared in sporadic stands (19 species in a single stand).
Community was distributed from loose sandy soil with low dunes to partially stabilized soil (with a cover of scattered stones) with mild undulates. Anabasis sp. was present on sand dunes and/or along dried watercourses having sandy soil. Other species appeared in different combinations under different distribution of high sand dunes or stabilized soil.
Zygophyllum sp. : Community represented an association of 21 species. Dominant cover was contributed by 0.3-1.0 m tall shrub of Zygophyllum sp., which was consistently associated with Pennisetum dichotomum and Rhazya stricta. Artemisia maritima, Withania coagulans, Haloxylon griffithii and Salsola arbuscula were though widely distributed yet only A. maritima contributed a significant cover. Other species were present in isolated stands. The community was present in relatively stabilized/loose slit soil. It was restricted to the hilly undulates, with watercourses having sharp edges. Vegetation was mainly present along dried watercourse. It occupied an extensive tract and was located at 600-1,000 m above sea level in southern Kharan (southern reaches of central highlands) and Rakshan valley (northern flanks of southern highlands).
Suaeda monoica: Community was located in southern highlands (central and eastern Khuzdar, alt. 600 m). It represented pure stands of S. monoica appearing in flat salt encrusted silt depressions.
Cocculus sp. : This community exhibited a reasonably uniform distribution of 5 species and a low vegetative cover. Cocculus sp. contributed major cover. Vegetation was mainly present along watercourses, which had gentle edges. Community was present in two closely running valleys, located at about 1,000 m above sea. These valleys represented gradually rising slopes with gentle undulations and loose silt soil.
Salsola foetida: Community presented an association of 6 species. It yielded a good cover of dried shrubs during winter. It was spread over a very wide tract of lowland valleys (at about sea level) of eastern Balochistan. The tract represented a flat plain of silty soil with patches of loose sand and scattered irrigated cultivation. S. foetida provided dominant cover with Suaeda ftuticosa working as second dominant. Out of 4 other associated species, Cynodon dactylon and Capparis decidua were better represented, while Ziziphus sp. and Calotropis sp. contributed a minor cover.
Capparis decidua: This community was distributed over an extensive area of central lowland plains (alt. <100 m) having silt soil but with scattered sandy patches. Community represented an association of 4 species yielding a low cover, mainly contributed by widely scattered large shrubs of C. decidua. Saccharum manja (scattered as bushes in depressions), Ziziphus sp. (isolated bushes or small trees) and Salsola foetida (along watercourses) were three other species present during winter.
Panicum antidotale: In central lowlands (at sea level) a low cover presented an association of 3 species spread over very wide tract in flat plains with hard clay soil. Slight depressions with very gentle slopes were scattered within the stand and were holding major part of vegetation. P. antidotale contributed the major part of cover, while scattered bushes of Salsola foetida and Calotropis sp. worked as second dominants.
Gymnocarpos fruticosum: Community was located in relatively stabilized sandy undulates appearing around bare depressions along seacoast (around Pasni, southern highlands). Total vegetative cover was very high (33.44 %) on undulates. G. firuticosum contributed the dominant cover in this association of 5 species. Rest of the cover is almost equally shared by other species, Riolirion montanum giving a very sparse appearance.
Scorzonera tortuosissima: It presented an association of 5 species yielding 12.92% cover. Thick vegetation was present in sandy patches, leaving rest of the plains with very sparse vegetation. It was located along coastline (west of Gwadar). General soil was loose sandy. Dominating species (S. tortuosissima) provided major part of absolute cover. Periploca aphylla, Pennisetum dichotomum, Hordeum vulgare and Heliotropium cabulicum shared rest of the cover.
Atriplex crassifolia: Community was present in lowland vast valley present in the extreme southwestern flanks of province. The stand had stabilized hard clay flats with low sandy undulations appearing in patches. The major part of cover was contributed by vegetation scattered on sand dunes. Community represented an association of 8 species. Major part of the cover is contributed by A. crassifolia. All other species had a minor contribution towards stand cover.
Convolvulus spinosus: In southern lowlands an association of 9 species exhibited a very low absolute cover, mainly contributed by very scattered bushes of C. spinosus. Rest of the cover is almost equally shared among 8 species, though Haloxylon salicornicum was better represented. Community was present in hilly undulates at 200 m above sea. Vegetation was scattered in depressions, having soil moisture and stabilization.
Cynodon dactylon: This community represented an association of 5 species distributed in extensive mild undulates or almost level plains located in southern lowlands along the coastline. Community was present in stabilized soil. C. dactylon bore dominant part of an overall good vegetative cover. Two other species (Lycium barbarum and an unidentified shrub, locally called as Mesk) contributed significant proportions, while Gymnocarpos ftuticosum and Citrullus colocynthis shared a minor part of a community cover.
Saccharum ciliare: An association of 10 species, located in southeastern lowlands at an altitude of 100-200 in represented this community. The dominant species (S. ciliare) contributed 62.24% of total cover. Euphorbia neriffiblia, Salvadora oleoides and Ziziphus jujuba had a scattered appearance. Soil was silt with scattered stone. Vegetation was evenly spread, though better cover was present in depressions.
Suaeda fruticosa: An association of 14 species, mainly distributed over a vast saline tract in coastal regions of Makran and Lasbella, located at about sea level, represented this community. Cynodon dactylon (3 stands), Capparis decidua (2) and the dominant species (so fruticosa) were present in more than one stand. The dominant species contributed some 71% of the cover, though C dactylon, Abutilon muticum, Launaea nudicaulis and C decidua also contributed a significant share. Vegetation was rich in sandy patches. A stand, representing an extensive tract in northern highland (alt. 1,700 m, Qila Saifullah-Zhob) also represented this community. The vegetative cover in this tract was, however, low.
Haloxylon salicornicum: This was an association of 12 species, appearing in different combinations in two stands having loose stabilized/sandy soil and saline character. A stand in northern highland hilly undulates (alt. 1,700 m) with widely scattered shrubs of 7 species yielded a very thin cover (1.17%). H. salicornicum contributed some 42% of relative cover and was associated with H. griffithii and Convolvulus microphyllus. Four other species (Peganum harmala, Alhagi camelorum, Suaedaftuticosa, Artemisia maritima) shared a minor cover. The other stand was present in southern lowlands (Makran, 100 m) bearing a better cover contributed by evenly scattered shrubs. Community, in this stand, represented other set of five species associated with H. salicornicum. Capparis decidua and Ziziphus sp. were scattered in favourable patches, while other smaller species were evenly distributed.
Phenology: The available data (Table 2) suggests that different broad localities give different phenological appearances during different parts of winter.
In northern highlands most of the species (74%) were dry during early winter. Foliage was present (16%) on Convolvulus microphyllus, Artemisia maritima and Stocksia brahuica, while limited flowers (11%) were present on Haloxylon griffithii and S. brahuica. During mid winter these valleys presented a very dry look (79%), though some green foliage (11%) persisted on S. brahuica and C. microphyllus, the former also bore fruits (6%). Chrysopogon serrulatus started sprouting towards the later part of mid winter. These valleys started giving a green look (90%) in late winter, especially in March-April, when all the species started sprouting and dry stalks constituted some 10% of the vegetation.
In central highlands a mixed phenological appearance was exhibited during early winter. One group of species (Calligonum comosum, Koeleria phleoides, Launaea glomerata, Stipa pennata, Cyperus niveus, Hyoscamus muticus, Centaurea moschata, Eragrostis cynosuroides, Pennisetum dichotomum, Cynodon dactylon, Alhagi camelorum) was represented by dry stocks (47%). The other group (Malcolmia afticana, M. bungai, Artemisia maritima, Peganum harmala, Euphorbia cheirolepis, Gaillonia aucheri, Haloxylon ammodendron, Heliotropium cabulicum, Nepeta glomerulosa, Saccharum munja, Anabasis sp., Tribulus alatus, Astragalus hyrcanus, Rhazya stricta, Citrullus colocynthis, Tamarix pallasii, Ziziphus sp.) had varying degree of foliage (3 1 %). Out of the second group some species (H. ammodendron, Anabasis sp., R. stricta, T pallasii) also bore flowers (16%) and/or (A. maritima, H. cabulicum, S. munja, C colocynthis, Ziziphus sp.) fruits (6%).
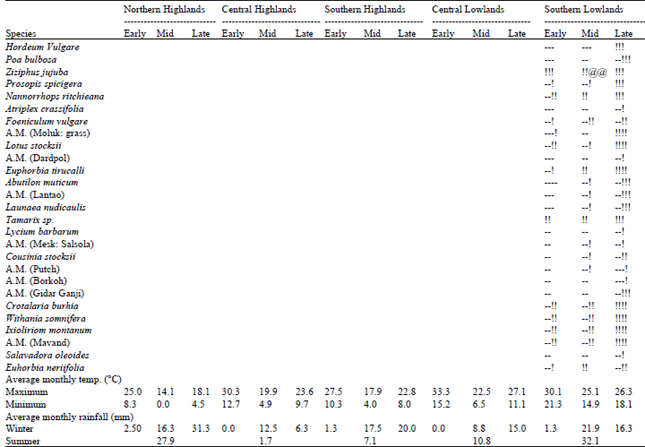
| Table 2: | Phenological variation exhibited by different plant species and associated data on temperature and rainfall in different broad localities of the Houbara wintering tracts of Balochistan (-= dry, != foliage, *= flowering @= seed/fruits; each symbol indicated 25% intensity) |
  | |
During mid winter, vegetation gave a drier look. Some of the species (S. arbuscula, H. cabulicum, T alatus, A. hyrcanus, Haloxylon salicornicum, Zygophyllum sp., N. glumerulosa) had completely dried (50%). Others (A. camelorum, P. dichotomum, E. cynisuroides, C niveus) started sprouting (30%) towards the end of mid winter, and C comosum also bore flowers (11%) and fruits (9%). Phenology of other species remained greatly unchanged from that of early winter. Late winter was characterised by sprouting of foliage (81%) on all the species, except for H. salicornicum and C moschata that remained dry (7%). Zygophyllum sp., T pallasii, Anabasis sp., T pallasii, Anabasis sp. and R. stricta bore flowers (7%) along with foliage, while fruits (5%) were present on Anabasis sp., C comosum and H. ammodendron.
In southern highlands, all the species gave a very dry look (78%) during early winter, except Anabasis sp. and Cocculus sp., which bore some degree of foliage (16%). Flowers were presents on 2% and fruits on 4% of the species. Dry vegetation (78%) persisted during mid winter, though Anabasis sp. started giving flowers (6%), fruits (2%) and some foliage (14%) was present on Ephedra intermedia. Most of the species were dry (40%) till late winter and sprouting (5 1 %), flowering (6%) and fruiting (3%) started towards the end of the winter.
Valleys in the central lowland were characterized by greener (52%) look during early winter. All the species were green except Saccharum ciliare, Andropogon sp., A. stocksil, Citrullus colocynthis, Ziziphus sp., Salsolafoetida, Cynodon dactylon, Calotropis sp., which were dry (17%). Capparis decidua bore fruits (13%). Flowers (19%) were present on C dactylon, S. foetida, Commiphora sp. and cultivated Eruca sativa. The general phenology remained unchanged till mid winter (38% foliage), however, C dactylon, C aphylla and S. foetida had' completely dried (41%), while P. harmala and Commiphora sp. bore fruits (18%). All the species had green foliage (78%) during late winter, while 22% of the vegetation was dry.
In southern lowlands, many species were green (31%) during early winter, except Periploca aphylla, Suaeda fruticosa, Scorzonera tortuosissima, Hordeum vulgare, G. ftuticosum, Poa bulbosa, A. crassifolia, L. nudicaulis, S. brahuica, L. barbarum and Cousinea stocksii, which were dry (63%). Flowers (2%) and fruits (4%) were present on some species. The phenology remained almost same during mid winter, except that some species (P. dichotoinum, C. spinosus, S. ciliare) were without green foliage (53%), while H. salicornicum, P. aphylla, G. ftuticosum, A. muticum, L. nudicaulis and C. stocksii started sprouting (42%). Fruits persisted on 5% of vegetation. During late winter, all the species (74%) gave a green look, while 22% of plants were dry stalks.
DISCUSSION
Very few comparable detailed phytosociological analysis are available for the general tract (Kayani, 1984; Kayani et al., 1984 and Marwat, 1988) and none from the Houbara habitat in Balochistan. Such studies undertaken along the global distribution range of Houbara (Collins, 1983, 1984 and Goriup, 1983b) also have a limited scope. The present study, therefore, is the first to present an elaborate data on phytosociological characters of area, but may be taken with care as these pertain to winters and to Houbara tracts only.
Floral diversity: The presence of at least 90 plant species, intra-stand homogeneity in the species composition, fewer species per stand, low constancy of appearance of majority of the species and recognition of 21 communities, when considered together suggest a wide diversity in composition of the phytohabitat adapted by Houbara. Such diversity in floral composition is expected in the wake of wide variation in temperature, altitude, rainfall and soil (Mian, 1983). The floral diversity in the area is expected to increase during early winter and early summer, with sprouting of ephemerals under favourable conditions of soil moisture and temperature, yet the wintering Houbara does not directly exploit it. This goes in an indirect conformity with many previous studies (Ali and Ripley,1971 and 1983; Roberts, 1991; Ponomareva, 1979; Alekseev, 1980; Taylor, 1983; Mian and Surahio, 1983; Karim and Hasan, 1983; Mirza, 1985 and Malik, 1985); each suggesting a different list of species for different areas of summering and wintering range of this race.
Stratification and sociability: All the stands share steppe desert vegetation. Shrubs constitute the only regular layer, herb and tree layer being irregular or appear in some part of the year. Vegetation exhibits varying degree of clumping. Major part of vegetation is limited to depressions, appearing between undulates and/or along watercourses. This pattern is typical of the deserts where depressions or edges of watercourses afford better soil moisture. Such a stratification/sociability pattern has been suggested previously as typical of Houbara habitat (Cramp and Simmons, 1980).
Vigour: A general vigorous look of all species and their propagation through sexual cycle (profuse flowering in Anabasis sp., Salsola arbuscula, Haloxylon ammodendron, Pteropyrum olivieri and fruiting on Citrullus colocynthis, Capparis decidua, Ziziphys sp., etc.) and/or vegetative propagation (vigorously sprouting in Calligonum sp., Stocksia brahuica, Artemisia maritima, Suaeda sp.) suggest that the plant species are fully adapted to prevailing conditions of temperature and rainfall and have adjusted their phenological cycles accordingly. The persistence of vigorous plants, especially in deeper deserts (which largely remain out of human interference), despite the severe and prolonged drought (spread over 3-4 consecutive years), may suggest a remarkable potential of vegetation to fight drought, which appears to hit the tract cyclically (Mian,1983 and Roberts, 1973, 1985). The vigorous state of the general plant species suggests a healthy general ecosystem.
Cover: All the communities and stands have a low vegetative cover. This can be attributed to arid character of the area (Ahmad, 1951; Ali, 1971; Emberger and lemee, 1962), density of vegetation being a direct character of precipitation (Walter, 1971 and Mond, 1954). Though no direct study is available from Houbara habitat in Balochistan, yet comparable studies on general phytoecology (Kayani and Sheikh, 1981 and Kayani et al., 1984) suggest that this is a general attribute of vegetation of this area. A similar low vegetative cover has been suggested by transect data available on Houbara habitat in Morocco (Goriup, 1983a).
Cover contributed by herbs is generally low. Major part of the herbs appears in isolated patches during spring and/or later summer. This goes in conformity with the transect data collected in later winter from Morocco (Goriup, 1983a). The major species listed for other wintering tracts of Pakistan also include very few annuals, except for those appearmg in cultivation as weeds (Goriup, 1983b; Taylor, 1983; Karim and Hasan, 1983; Mirza, 1985 and Malik, 1985).
Communities: The recognition of communities on absolute vegetative cover seems justified. This is because during winter vegetative cover is mainly contributed by shrubs of an almost equal height and reasonably identical sociability. The absolute cover, under these conditions, is a direct indicator of the relative cover, frequency and density, the three parameters generally exploited for calculation of importance value and thence to work out the dominant species (Curtis and McIntosh, 1950). Recognition of plant communities, in strict phyto-sociological sense may not be of a direct value in studies on animal habitat utilization and has not been attempted in allied previous studies on bustards (Goriup, 1983a; Collins, 1983, 1984; de Trucios and Almansa, 1989).
Different communities are distributed in accordance with ecological amplitude of the dominant species. In northern highlands isolated stands representing Suaeda ftuticosa (stabilized saline valleys), Andropogon halepensis (loose sandy good soil moisture), Stocksia brahuica (stabilized slope soil), Haloxylon griffithii (stabilised hilly undulates) and Haloxylon salicornicum (loose sandy hilly undulates with high salts) communities are disturbed in isolated tracts under different soil conditions. In central highlands, Haloxylon ammodendron and Anabasis sp. communities run side by side, the former in high sand dunes and the later in low sand dunes. Zygophyllum sp. and Cocculus sp. communities are present in stabilized soil, the former in the hilly undulates and later in slope valleys. A pure stand of Rhazya stricta is present in hilly flats with stabilized sandy soil. In southern highlands, Suaeda monoica community is present in salt encrusted plains. Astragalus hyrcanus (stabilized sand in hilly undulates), Rhazya stricta (stabilized silt hilly undulates), Haloxylon griffithii (silt hilly valleys) and Anabasis sp. (low sandy undulates) communities are distributed in varying degrees of stabilized soil and undulations.
Central lowlands (at almost sea level) have Salsolafoetida (stabilized silt soil), Capparis decidua (stabilized sandy soil) and Panicum antidotale (plains having hard clay soil) communities in different stretches. In southern lowlands, Suaeda ftuticosum community is widely distributed in saline sandy coastal tracts and Gymnocarpos ftuticosum in relatively stabilized coastal dunes. The other communities including, Scorzonera tortuosissima, Convolvulus spinosus, Atriplex crassifolia, Cynodon dactylon and Saccharum ciliare are distributed at relatively higher altitudes and in hilly valleys having varying degrees of sandy/silt soil.
Phenology: The majority of desert-adapted plants reproduce vegetatively and through fruits and seeds. Rainfall in a year or part of the year can ensue sprouting of vegetative parts, depending upon suitability of temperature. Degree of green foliage. in an area is decided to a considerable degree by rainfall gradient/pattem (Walter, 1954). Flowering is basically controlled by photoperiods. Therefore, flowering/fruiting appear during specific part of the year. Suitability of temperature, humidity and soil moisture contributes positively towards intensity of flowering and seed production/germination. The general reliance of the species on both vegetative propagation and seed production/germination attributes a higher survival value and ensures their survival under all odds.
Phenological cycle followed in different broad localities can be explained on temperature and rainfall patterns. Under the influence of better rainfall and mild temperature during summer in northern highlands, the major part of the flora bear fruits/seeds towards the end of summers. The low temperature forces these species to remain dormant during mid winter. The seeds/vegetative parts sprout towards late winter or early summer, under winter rains and favourable temperature. In central highlands, the harsh summer temperature associated with limited rain results in very dry summer vegetation. Mild temperature during early winter causes some species to bloom while the others bear green foliage. The quantum of green foliage decreases during mid winter under low temperature. Sprouting in all the species occurs during late winter under moderate temperature and winter rain. In contrast, better summer rains, mild winter temperature and reasonable winter rains in central lowlands provide green foliage throughout the winter with major species flowering during late summer or early winter. The southern highlands, with limited summer rain and harsh winter temperature, give a rather dry look till mid winter and leaves, flowers and/or fruits appear in late winter. The southern lowlands receive more liberal summer rains with some winter rains and bear a comparatively mild winter temperature. The plant species in these tracts bear green foliage during winter, most of the species flowering/fruiting during summer.
Grazing stress: A higher proportion of perennials suggest that, in general, the grazing stress remain within permissible limits as suggested by Dabadghoa and Shankarnarayan (1973). This is understandable according to nomadic pattern of life followed till recent past. Under such pattern grazing stress, coming from livestock, is shared between different tracts, especially during unfavourable seasons and/or years. The development of permanent human settlements in the' area can pose a future threat to the vegetation of the area and bare patches have already started appearing around such settlements (Mian, 1987).
ACKNOWLEDGMENTS
The study was conducted with a partial financial support of WWF-Pakistan and field support of Balochistan Forest Department. Thanks are also due to a number of field workers for their help during field studies.
REFERENCES
- Mueller-Dombois, D. and H. Ellenberg, 1974. Aims and Methods of Vegetation Ecology. 1st Edn., John Wiley and Sons, New York, USA., ISBN-13: 978-0471622901, Pages: 570.
Direct Link