
Ram P. Yadav
Natural Product Laboratory, Department of Zoology, D.D.U, Gorakhpur University, Gorakhpur- 273 009 (U.P.), India
Digvijay Singh
Natural Product Laboratory, Department of Zoology, D.D.U, Gorakhpur University, Gorakhpur- 273 009 (U.P.), India
S. K. Singh
Natural Product Laboratory, Department of Zoology, D.D.U, Gorakhpur University, Gorakhpur- 273 009 (U.P.), India
Ajay Singh
Natural Product Laboratory, Department of Zoology, D.D.U, Gorakhpur University, Gorakhpur- 273 009 (U.P.), India
Pakistan Journal of Biological Sciences
Year: 2003 | Volume: 6 | Issue: 14 | Page No.: 1223-1228







ABSTRACT
The exposure to fish over 96 h to sub-lethal doses 8.24 mg, DW L-1 and 16.48 mg, DW L -1 of stem-bark extracts of Croton tiglium of family Euphorbiaceae significantly altered the level of total protein, total free amino acids, nucleic acids, pyruvate, lactate, glycogen and activity of succinic dehydrogenase, lactic dehydrogenase, cytochrome oxidase, acetylcholinesterase, acid and alkaline phosphatase and protease activity in muscle, liver and gonadal tissue of fish Channa punctatus. The alteration in all the biochemical parameter was significantly dose dependent. Withdrawal study also shows that there is a partial recovery in the levels of glycogen, pyruvate, nucleic acids, acetylcholinesterase, cytochrome oxidase and acid and alkaline phosphatase, but nearly complete recovery in total protein, total free amino acids level and activity of protease, lactic dehydrogenase, succinic dehydrogenase and lactate in all the three tissues of the fish after the 7th day of the withdrawal of treatment, which supports the view that the plant product is safer in use as pesticides for control of common weed fishes in culture ponds.
PDF Abstract XML References Citation
How to cite this article
Ram P. Yadav, Digvijay Singh, S. K. Singh and Ajay Singh, 2003. Metabolic Changes in Freshwater Fish Channa punctatus Due to Stem-bark Extract of Croton tiglium. Pakistan Journal of Biological Sciences, 6: 1223-1228.
DOI: 10.3923/pjbs.2003.1223.1228
URL: https://scialert.net/abstract/?doi=pjbs.2003.1223.1228
DOI: 10.3923/pjbs.2003.1223.1228
URL: https://scialert.net/abstract/?doi=pjbs.2003.1223.1228
Introduction
The ultimately use of synthetic pesticides and large amount of fertilizers as a pond treatment have not only affected the freshwater ecosystem adversely but also the immunological disorder of the freshwater animals (Richard et al., 1991). In recent years, the use of medicinal plants as effective alternatives of synthetic pesticides and fertilizers has gained importance especially to combat problem both in fish and aquatic environment (Dahiya et al., 2000), because they are highly toxic to the target pests (Marston and Hostetmann, 1987; Singh and Agarwal, 1988; Singh et al., 1996; Yadav and Singh, 2001).
The plant Croton tiglium (Family-Euphorbiaceae) is common medicinal plant of India having high molluscicidal properties (Yadav and Singh, 2001). The present study was to know inducted the mode of action and long-term effect of these plant products on non-target animals, because these substances cannot be put to commercial use without a study of these aspects as well.
MATERIALS AND METHODS
The fish Channa punctatus (Bloch) commonly called snake headed fish (17.5±1.20 cm), was collected Ramgarh lake of Gorakhpur district. The collected animals were stored in glass aquaria containing 100 L of dechlorinated tap water. Prior to the experiment, the fishes were allowed to acclimate to laboratory conditions for 7 days. Diseased, injured and dead fish (if any) were removed as soon as possible to prevent the decomposition of the body. Water was changed every 24 h. Average-sized adult animals were used for the experiment.
Stem-bark of Croton tiglium (Family-Euphorbiaceae) were collected from Botanical garden of D.D.U, Gorakhpur University Gorakhpur and identified by Prof. S.K. Singh (taxonomist) Department of Botany D.D.U, Gorakhpur University, Gorakhpur.
Preparation of aqueous extracts of stem bark: The dried powder (10 mg mL-1) were minced with distilled water homogenized for 5 min and centrifuged at 1000 g for 10 min, the supernatant was used a water extract for the further studies.
The acclimatized fishes were treated with sub-lethal doses (8.24 mg, DW L-1 and 16.48 mg, DW L-1) of the stem-bark of the plant Croton tiglium for 96 h exposure periods according to the method of Singh and Agarwal (1988). Six aquaria were set up for each dose and each aquarium contained 10 fishes in 6 L of dechlorinated tap water The LC50 of stem-bark extracts of Croton tiglium against snail was 20.60 mg l-1 and 6.11 mg l-1 for 24 and 96 h, respectively (Yadav and Singh, 2001).
During exposure to sub-lethal doses, observations were made to detect the physical symptoms of poisoning in the fish. Behaviour symptoms like the fishes starting to experience respiratory distress, gulping of air at the upper surface ere observed in the beginning of the experiment.
In order to see the effect of withdrawal of the extract treatment, the fishes were exposed to 96 h to 16.48 mg, DW L-1 and the one half of the animal was sacrificed and the level of total protein, total free amino acids, DNA, RNA and glycogen was measured. The other half was transferred to stem-bark free water, which was damaged every 24 h for the next 6 days, following this, the level of total protein, total free amino acid, DNA, RNA, glycogen, pyruvate, lactate and activity of acetylcholinesterase, succinic dehydrogenase, lactic dehydrogenase and acid and alkaline phosphatase was measured in muscle, liver and gonadal tissue of the fishes. Control animals were kept in dechlorinated tap water without any treatment.
Each assay was replicated six times and values (mean±SE) have been expressed. Students’ ‘t’ test was applied to locate significant (P<0.05) differences between treated and control animals (Sokal and Rohlf, 1973).
Protein: Protein levels were estimated according to the method of Lowry et al. (1951) using bovine serum albumin as standard. Homogenates (5 mg ml-1, w v-1) were prepared in 10%TCA. Values have been expressed as μg mg -1 of tissue.
Total free amino acids: Estimation of total free amino acid was made according to the method of Spices (1957). Homogenates (10 mg ml-1, w v-1) were prepared in 95% ethanol, centrifuged at 6000 xg and used for amino acid estimation. Free amino acids have been expressed as μg mg-1 of tissue.
Nucleic acids: Estimation of DNA and RNA was performed, by methods of Schneider (1957) using diphenylamine and orcinol reagents, respectively. Homogenates (1 mg ml-1, w v-1) were prepared in 5% TCA at 900C, centrifuged at 5000 g for 20 min and supernatant was used for estimation. Both DNA and RNA have been expressed as μg mg-1 of tissue.
Glycogen: Glycogen was estimated by the Anthrone method of Van Der Vies (1954). In present experiment 50 mg of tissue was homogenised with 5 ml of cold 5%TCA. The homogenate were filtered and 1.0 ml of filtrate was used for assay. Results have been expressed as mg g -1 of tissue.
Pyruvate: Pyruvate level was measured according to Friedemann and Haugen (1943). Homogenate (50 mg ml-1, w v-1) was prepared in 10% TCA. Sodium pyruvate was taken as standard. Result has been expressed as μg pyruvate mg-1 of tissue.
Lactate: Lactate was estimated according to Barker and Summerson (1941), modified by Huckabee (1961). Homogenate (50 mg ml-1, w v-1) was prepared in 10% cold TCA. Sodium lactate was taken as standard. Lactate level has been expressed as μg mg-1 of tissue.
Protease: Protease activity was estimated by the method of Moore and Stein (1954). Homogenate (50 mg ml-1, w v-1) was prepared in cold distilled water. Optical density was measured at 570 nm. The enzyme activity was expressed in μ mol of tyrosine equivalent mg-1 protein h-1.
Acid and alkaline phosphatase: Activities of acid and alkaline phosphatase were measured by the method of Bergmeyer (1967) and modified by Singh and Agarwal, (1982). Tissue homogenate (2% w/v) were prepared in ice cold 0.9% saline and centrifuged at 5000 xg at 0°C for 15 min. Optical density was measured at 420 nm against a blank, prepared simultaneously. The enzyme activity has been expressed as amount of ρ-nitrophenol formed -1 30 min mg -1 protein in supernatant.
Lactic dehydrogenase: Lactic dehydrogenase (LDH) activity was measured according to the method of Anonymous (1984). Homogenates (50 mg ml-1, w v-1) were prepared in 1 ml of 0.1 M phosphate buffer, pH 7.5 for 5 min in an ice bath. Enzyme activity has been expressed as nanomol of pyruvate reduced-1 min mg-1 protein.
Succinic dehydrogenase: Succinic dehydrogenase activity was measured by the method of Arrigoni and Singer (1962). Homogenate (50 mg ml-1, w v-1) was prepared in 1 ml of 0.5M potassium phosphate buffer, pH 7.6 for 5 min in an ice bath. Optical density was measured at 600 nm. Enzyme activity has been expressed as μmol dye reduced-1 min mg-1 protein.
Cytochrome oxidase: Cytochrome oxidase activity was measured according to the method of Cooperstein and Lazarow (1951). Homogenates (50 mg ml-1, w v-1) were prepared in 1 ml of 0.33 M phosphate buffer (pH 7.4) for 5 min in ice bath. Enzyme activity has been expressed in arbitrary units-1 min mg-1 of proteins.
Acetylcholinesterase: Acetylcholinesterase was estimated by the method of Ellman et al. (1961). Homogenates (50 mg ml-1, w v-1) were prepared in 0.1 M phosphate buffer in ice bath. Optical density was measured at 412 nm at 25°C. Enzyme activity expressed in μmol ‘SH’ hydrolysed min mg-1 protein.
RESULTS
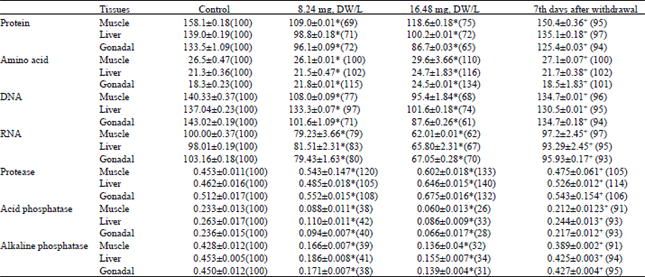
Data indicates a significant (P<0.05) dose-dependent decrease in the protein level in the muscle, liver and gonadal tissue of fish Channa punctatus exposed to sub-lethal doses (8.24 mg, DW L-1 and 16.48 mg, DW L-1) of the stem-bark of Croton tiglium. Exposure to sub-lethal doses 8.24 mg, DW L-1 and 16.48 mg, DW L-1 of 24 h LC50 of stem-bark extracts resulted in a decrease of the protein level in the muscle to 69 and 75%, in the liver tissue to 71 and 72%, while in the gonads to 72 and 65% of the control value, respectively (Table 1). Exposure to 8.24 mg, DW L-1 and 16.48 mg, DW L-1 of the stem-bark extracts up to 96 h resulted in an increase of free amino acids level in the muscle to 100 and 110%, in the liver to 102 and 116%, respectively, while in the gonads to 115 and 134% of the control, on exposure to 8.24 mg, DW L-1 and 16.48 mg, DW L-1 respectively. The sub-lethal doses resulted in significant decline in nucleic acid level (DNA and RNA) in the muscle, liver and gonadal tissue of the fish. In the case of DNA level, decrease in the muscle to 77 and 68%, in the liver to 97 and 74%, while in the gonadal tissue to 71 and 61% of control, respectively, were observed, and the same trend was observed in the RNA level, the decrease noted to 79 and 62% in the muscle, 83 and 67% in the liver and 80 and 70% in the gonads of the control, respectively (Table 1). The sub-lethal exposure to stem-bark extracts of Croton tiglium, caused a significant reduction in the muscle, liver and gonadal content of the fish (Table 1).
The 96 h exposure to 8.24 mg, DW L -1 and 16.48 mg, DW L-1 of the stem-bark caused a decrease in muscle glycogen content to 70 and 65%, in the liver to 67 and 63%, while in gonads to 68 and 59% of the control respectively. The 96 h exposure to sub-lethal doses of 8.24 mg, DW L-1 and 16.48 mg, DW L-1 of the stem-bark extracts caused a significant decrease in the muscle pyruvate level 48 and 30% and 58 and 35% in liver, while in the gonads to 46 and 28% of control, respectively. The 96h exposure to sub-lethal doses 8.24 mg, DW L-1 and 16.48 mg, DW L-1 of stem-bark extracts caused a increase in the muscle lactate level to 110, 140 and 121, 147% respectively, while in the gonads to 108 and 137% of the control (Table 2).
The 96 h exposure to sub-lethal doses 8.24 mg, DW L-1 and 16.48 mg, DW L-1 of stem-bark extracts caused a lactic dehydrogenase (LDH) activity was reduced to 88, 80% in muscle and 89%, 77% in liver and while 91 and 77% in gonads of controls, respectively (Table 2). Activity of the cytochrome oxidase was reduced to 81%, 70% in muscle and 80 and 68% in liver and 83 and 71% of the fish of gonads respectively. Acetylcholinesterase activity was reduced to 50 and 28% in muscle and 55, 36% in liver, while 52 and 28% in gonadal of fish, respectively. The succinic dehydrogenase activity was increased to 107, 118% in muscle and 105 and 120% in liver and 109 and 116% of controls after 96 h treatment to the sub-lethal doses of 8.24 mg, DW L-1 and 16.48 mg, DW L-1, respectively (Table 2).
| Table 1: | Changes in total protein, total free amino acids, nucleic acid (DNA and RNA) (μg mg-1) level and activity of protease (μmol of tyrosine equivalents mg-1 protein h-1) and acid and alkaline phosphatase (μmol substrate hydrolysed 30 min mg-1 protein) in different tissues of freshwater fish Channa punctatus after 96 h exposure to sub-lethal doses (8.24 mg, DW L-1 and 16.48 mg, DW L-1) of stem-bark extracts of Croton tiglium and 7th days after withdrawal |
 | |
Values are mean ±SE of six replicates. Values in parentheses are % level with control taken as 100%. Data were analyzed through student’s ‘t ’test. *, Significant (P<0.05), when treated groups were compared with control +, Significant (P<0.05) when students ‘t’ test was applied between treated (16.48 mg, DW LG1) and withdrawal groups. | |
| Table 2: | Changes in glycogen (mg g-1), pyruvate (μmol g-1), lactate (mg g-1) level and activity of LDH (μmol mg-1 protein h-1), SDH (μmol of dye reduced min-1 mg protein-1), cytochrome oxidase (arbitrary unit min-1 mg protein-1) and AChE (μmol ‘SH’ hydrolysed min-1 mg protein-1) in different tissues of Channa punctatus after 96 h exposure to sub-lethal doses (8.24 mg, DW L-1 and 16.48 mg, DW L-1) of stem-bark extracts of Croton tiglium and 7th days after withdrawal |
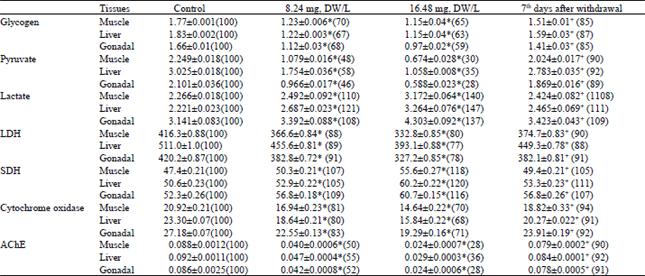 | |
The acid phosphatase activity was reduced to 38%, 26% in muscle and 42 and 33% in liver, while 40 and 28% in gonadal of fish, respectively. The alkaline phosphatase activity was reduced to 39%, 32% in muscle and 41 and 34% in liver and 38 and 31% of controls after 96 h treatment to the sub-lethal doses of 8.24 mg, DW L-1 and 16.48 mg, DW L-1 respectively (Table 1). The protease activity was increased to 120 and 133% in muscle and 105 and 140% in liver, while 108 and 132% in gonadal of fish, respectively (Table 1).
DISCUSSION
It is evident from the results presented here that the extracts of stem-bark of Croton tiglium besides being potent molluscicides (Yadav and Singh, 2001) are toxic to fish Channa punctatus at higher concentrations and longer exposure periods. It is also clear from the present study that the treatment with sub-lethal doses of 8.24 mg, DW L-1 and 16.48 mg, DW L-1 contained up to 96 h significantly altered the levels of total protein, total free amino acids, nucleic acids, glycogen, pyruvate and lactate level and activity of protease, lactic dehydrogenase, acetylcholinesterase, succinic dehydrogenase, acid and alkaline phosphatase and cytochrome oxidase. The rate of alteration in all the cases was significantly (P<0.05) dose-dependent.
The protein is the energy source to spore during chronic period of stress. Animal exposed to sub-lethal concentration of toxicant experienced, the greater stress during the process of detoxification of given toxicant on the metabolic level of animal. The metabolic rate of animal reared at different concentration of toxicant was greater than that of animal reared in freshwater (Aranachalem et al., 1980). The protein depletion in treated animal was the physiological strategy played by the animal to adopt itself to the changed metabolic systems. This leads to degradative processes like proteolysis and utilization of degraded products for increased metabolism.
The decrease in protein level observed in present investigation may be due to their degradation and also to their possible utilization for metabolic purpose. Bradbury et al. (1987) pointed out that the decreased protein content might also be attributed to the destruction or necrosis of cells and consequent impairment in protein synthesis machinery. The quantity of protein is dependent on the rate of protein synthesis, or on the rate of its degradation. The quantity of protein may also be affected due to impaired incorporation of amino acids in to polypeptide chains (Singh et al., 1996).
The accumulation of free amino acids can also be attributed to lesser use of amino acids (Seshagirri et al., 1987) and their involvement in the maintenance of an acid-base balance (Moorthy et al., 1984). It has been suggested that the stress condition in general induce elevation in the transamination pathway (Natrajan, 1985). It is supported by the observation of Malla Reddy and Bashamohiden (1995). Since, euphorbious plant is used in this study may also have same antiphosphatase activity. So the reduction in protein level may be due to the inhibition of alkaline phosphatase activity, as it plays an important role in protein synthesis (Pilo et al., 1972) and other secretary activities (Ibrahim et al., 1974).
The decrease in cytochrome oxidase might be either the result of reduced availability of O2, which in turns has reduce the capacity of electron transport system to produce ATP molecules or should be due to the direct impact of the active moiety reactions like the function of the cytochrome oxidase in electron transport system. Since Euphorbiales are anticholinesterase inhibitor, their poisoning effecting Kreb’s cycle thereby diminishing the rate of electron transport system and oxidative phosphorylation, resulting in less synthesis of ATP.
Reduction of glycogen level supposes to be the result of greater stress, the organs experienced during the process of detoxification of active moieties and their metabolites. Anti-acetylcholinestrase agents are known to inhibit acetylchlinesterase in nerve and other tissue cause an increase in acetylcholic content after intoxication (Kaundinya and Ramamurthy, 1978). Nilsson et al. (1976) reported and increase in Ach content has been shown to enhance the secretion of catecholamines in cod, Gardus morhua, which may bring about glucogenolysis and hyperglyceimia through the reserved level of cyclic AMP (Terrior and Perrier, 1975).
In the present study, the glycogenolysis seems to be the result of increased secretion of catecholamines due to stress of plant extracts treatment, because the latices of euphorbious plants have potent anti-AchE activity against snails (Singh and Agarwal, 1991). According to Umminger (1977) reported carbohydrate represents the principal and immediate energy source for animals exposed to stress conditions carbohydrate reserve depleted to meet energy demand. The major raw materials of gluconeogenesis are lactate and alanine produced by active and erythrocytes. The rate of production of pyruvate by glycolysis exceeds the rate of oxidation of pyruvate by the citric acid cycle (Sambasiva Rao, 1999).
Lactate is a dead end in metabolism. It must be converted back in to pyruvate before it can be metabolised. The only purpose of the reduction of pyruvate to lactate is to regenerate NAD+ so that glycolysis can proceed in active skeletal muscle and erythrocytes. Note that the conversion of glucose to lactate does not involve a net oxidation-reduction. Rather, one of the carbon atoms in lactate is more oxidized than in glucose and another is more reduced (Siva Prasada, 1980).
Withdrawal experiments were performed to see whether, biochemical alteration by exposure to plant moiety would return normal if the treatment were discontinued. There was nearly complete recovery of total protein, total free amino acid, lactate, cytochrome oxidase, SDH, protease and a partial recovery of glycogen, nucleic acid (DNA and RNA), pyruvate, LDH, AChE and acid and alkaline phosphatase activity in the different body tissues of fishes.
We therefore believes that these plant extracts may eventually be of great values for the control of aquatic target organisms as well as predatory fishes.
ACKNOWLEDGMENTS
One of the authors (Ram P. Yadav) is thankful to Department of Environment and Forest Govt. of India (Sanction No. F-14/35/96/ MAB-RE dated 9.11. 1999) for financial assistance.
REFERENCES
- Arunachalam, S., K. Jeyalakshmi and S. Aboobucker, 1980. Toxic and sub-lethal effects of carbaryl on a freshwater cat fish Mystus vittatus. Arch. Environm. Cont. Toxicol., 9: 307-316.
CrossRef - Ellman, G.L., K.D. Courtney, V. Andres Jr. and R.M. Featherstone, 1961. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol., 7: 88-90.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Natarajan, G.M., 1985. Induction of branchial enzymes in snake head (Channa striatus) by oxydemeton-methyl. Pestic. Biochem. Physiol., 23: 41-46.
CrossRefDirect Link - Singh, A. and R.A. Agarwal, 1988. Possibility of using latex of Euphorbiales for snail control. Sci. Total Environ., 77: 231-236.
CrossRef - Singh, A., D.K. Singh, T.N. Misra and R.A. Agarwal, 1996. Molluscicides of plant origin. Biol. Agric. Horticult., 13: 205-252.
CrossRef - Yadav, R.P. and A. Singh, 2001. Environmentally safe molluscicides from two common euphorbiales. Iberus, 19: 65-73.
Direct Link