
Zaheer Ahmad Nasir
Botany Department, Government, F.C. College, Lahore-54600, Pakistan
Sikander Sultan
Botany Department, Government, F.C. College, Lahore-54600, Pakistan
Pakistan Journal of Biological Sciences
Year: 2002 | Volume: 5 | Issue: 7 | Page No.: 758-762







ABSTRACT
A total of 78 weed species were recorded within gram, lentil, mustard and wheat crops. They were distributed among 28 families, including 2 monocots. The most important families were Asteraceae, Poaceae (11 spp. each), Papilionaceae (7 spp.), Boraginaceae (6 spp.), Amaranthaceae (5 spp.) and Solanaceae (4 spp.), while the remaining families had less than four species. There were 47 weed species in gram fields, 41 in lentil fields, 53 in mustard fields and 59 in wheat fields. Maximum contribution was made by therophytes (49 spp.) followed by hemicryptophytes (18 spp.) and chamaephytes (10 spp.). One parasitic weed species was also recorded. Except microphyllous (12 spp.) and mesophyllous (3 spp.), most of the species were leptophyllous and nanophyllous with 26 and 37 species, respectively. Leaf size of most weeds were reduced indicating an adaptation to arid region. A quantitative analysis is required to determine the ecological status of these weeds in various crops for their proper management.
PDF Abstract XML References Citation
How to cite this article
Zaheer Ahmad Nasir and Sikander Sultan, 2002. Floristic, Biological and Leaf Size Spectra of Weeds in Gram, Lentil, Mustard and Wheat Fields of District Chakwal, Pakistan. Pakistan Journal of Biological Sciences, 5: 758-762.
DOI: 10.3923/pjbs.2002.758.762
URL: https://scialert.net/abstract/?doi=pjbs.2002.758.762
DOI: 10.3923/pjbs.2002.758.762
URL: https://scialert.net/abstract/?doi=pjbs.2002.758.762
INTRODUCTION
Weeds are natural hazards to the interests and activities of man (Mortimer, 1990). These are those plant species which damage the yielding potential of the land surface or water volume managed by man (FAO, 1994). Weeds compete with the crop plants for nutrients, water and sunlight and cause yield losses to a varying degree depending upon their species and population (Nayyar et al., 1994).
Due to these facts the removal and control of weeds become of prime importance in crops. However, without knowing the weeds, one cannot take steps in the right direction (Ayaz et al., 1995). The collection and identification of weeds have always been a pre-requisite for any ecological approach made towards the eradication and control of weeds. As such floristic composition and identification of weeds must be worked out (Murad et al., 1995).
The weed flora of different agricultural crops from various parts of Pakistan have been reported. The weed flora of tobacco fields (Hussain et al., 1986), sugar beet (Hussain and Rashid, 1986), cotton (Malik et al., 1988), turmeric (Rashid et al., 1992), rice and maize (Subhan et al., 1985; Malik and Hussain, 1990a; Siddiqui et al., 1992; Afzal et al., 1994), sugar cane (Irshad et al., 1980), potato (Murad et al., 1995) and wheat (Bokhari et al., 1986; Hussain et al., 1988; Shinwari et al., 1990; Ahmad, 1992; Ayaz et al., 1995; Hussain et al., 1998; Qureshi and Bhatti, 2001) have been reported.
Life form is primarily determined by heredity selection, it may be regarded as an adjustment of the vegetative plant body and life history to the habitat (Dastagir et al., 2000). Leaf spectra determines the growth and adaptational behaviour of species. Very little work is available on this aspect (Rashid et al., 1988; Hussain et al., 1985,1987,1993; Ayaz et al., 1993; Murad et al., 1995).
However, there is no such information available on the weed flora of gram, lentil, mustard and wheat fields from any part of the district Chakwal, so the present study was undertaken to report the weed flora from gram, lentil, mustard and wheat fields of the district Chakwal. These findings might be of great help to weed ecologists, agronomists and other agriculture scientists involved in their management.
MATERIALS AND METHODS
District Chakwal lies on the main historic route from old Taxila to Jehlum. It comprises of three tehsils named: Chakwal, Tala Gang and Choa Saiden Shah. Of the total 16,52,000 acres area of the district, 8,20,000 acres is cultivated (Anonymous, 2000).
Twelve sub-localities within three tehsils of the district Chakwal namely: 1. Bhaun, 2. Kalar Kahar, 3. Bochal, 4. Dudyal, 5. Pindi Gugran, 6. Balkasar, 7. Kot Sarang, 8. Jahtla, 9. Taman, 10. Puchnand, 11. Dulmyal and 12. Ratucha, where gram, lentil, mustard and wheat are grown, periodically visited for collection of weeds throughout the growing season. The collected weed plants were dried and mounted on the herbarium sheets. The weeds were then identified with the help of available literature (Nasir and Ali, 1994; Stewart, 1972) and also by comparison with herbarium specimens in NARC, Islamabad. Weeds were classified into various life forms and leaf size classes following Hussain (1989) and Raunkiaer (1934). Biological spectrum was also determined. The study covers a period from October, 99 to April 2000.
RESULTS AND DISCUSSION
A total of 78 species were recognized as weeds in gram, lentil, mustard and wheat fields of district Chakwal. Among them 66 species, distributed in 26 families, belonged to dicotyledons, while 12 species represented monocotyledons (Table 1).
The families in dwindling order of weed species were Asteraceae, Poaceae (11 spp. each) Papilionaceae (7 Spp..), Boraginaceae (6 spp..), Amaranthaceae (5 spp.), Solanaceae (4 spp.), Chenopodiaceae, Caryophyllaceae, Euphorbiaceae and Brassicaceae (each with 3 spp.). The remaining families had less than 3 species. The greater number of species in Asteraceae and Poaceae might be due to the excellent vegetative growth behaviour, fabulous seed production, efficient dispersal mechanism, long lasting dormancy and spectacular competitive ability of weed species present in these families.
Fruits of family Asteracea possess different types of pappus which help in their efficient dispersal. In the same way, members of family Poaceae produce seeds of extremely small size, which blow to long distances by wind thus causing an enormous increase in their population growth. Holm et al. (1977) reported Poaceae and Asteraceae as families having maximum number of weed species i.e., 44 and 32, respectively.
In all, 47 weed species were recorded from gram fields. Of them, Asphodelus tenuifolius Cavan., Carthamus oxycantha M.B., Convolvulus arvensis L., Digera muricata (L.) Mart., Sorghum halepense (L.) Pers. and Vicia monantha L. were most common. The prevalence of these weeds might be due to their high physiological efficiency which help them to absorb nutrients faster, utilize water better and accumulate dry matter at a more rapid rate.
Lentil fields had 41 species. Lathyrus sativus L., Euphorbia helioscopia L., Fumaria indica (Hausskn.) H.N., A. tenuifolius Cavan. and Calendula arvensis L. were most prevalent among them. A root parasite Orobanche aegyptiaca Pers. was also recorded.
| Table 1: | Floristic list, life form and leaf size spectra of weeds in gram, lentil, mustard and wheat crops of district Chakwal during 1999-2000. |
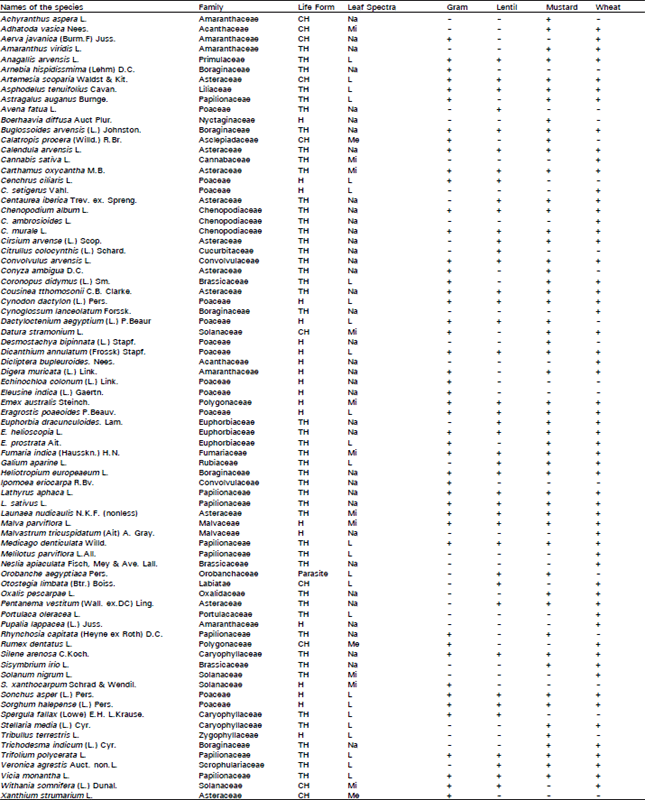 | |
| Key: TH = Therophytes, CH = Chamaephytes, H = Hemicryptophytes, P = Parasite, L = Leptophyll, Na = Nanophyll, Mi =Microphyll, Me = Mesophyll, + = Present, – = absent. | |
| Table 2: | Biological spectrum of weeds in the gram, lentil, mustard and wheat fields of district Chakwal. |
 | |
| Table 3: | Leaf size spectra of weeds in the gram, lentil, mustard and wheat fields of district Chakwal. |
 | |
The copiousness and success of these weeds might be due to their large seed bank in the soil and ability to disperse, particularly in the disturbed environment. Cropping pattern and cultural practices of the last few years controls which species of weed flowered and contributed to weed seed bank in soil (Anonymous, 1987).
Weed flora of mustard fields comprised of 53 species. The most frequent weeds were Artemesia scoparia Waldest and Kit., Calendula arvensis L., C. oxycantha M.B., Convolvulus arvensis L., F. indica (Hausskh.) H.N., Lathyrus aphaca L. and S. halepense L. Pers. One parasitic weed O. aegyptiaca was also collected from mustard fields. High reproductive potential might be the possible reason of abundance of these weeds. According to Joshi and Nigam (1970) seeds of some weeds have high reproductive capacity and great recycling potential.
Among the 59 weed species in wheat fields Chenopodium album L., C. murale L., A. tenuifolius Cavan., Anagallis arvensis L., Astragalus augans Burnge., C. oxycantha M.B., Cirsium arvense (L.) Scop., Convolvulus arvensis L. and F. indica (Hausskn.) H.N. were the most common. Variation in germination behaviour, discontinuous and internally controlled germination and great longevity of seeds in soil might be the prime factors which ensures their emergence and spread in a particular agroecosystem. Seed dormancy reflects a response to adverse conditions and inevitably leads to persistent seed banks as opposed to transient ones (Grime, 1989).
According to Hanson and Churchill (1965) floristic lists are valuable for characterization. It had been observed that weeds differ in their time of germination. A. arvensis L., A. scoparia Waldest and Kit., A. tenuifolius Cavan., Calendula arvensis L., C. oxycantha M.B., C. album L., C. murale L. and Convolvulus arvensis were recorded in all the crops throughout the season. According to Hussain and Malik (1986) a weed may be regularly recorded in many crops providing their ecological range and phenological cycle overlap with each other. These species owe their uniform presence due to high seed out put and thus high capacity to colonize, perpetuate and establish themselves (Malik et al., 1988).
Some weeds were collected in late season due to their late germination, i.e. Silene arenosa C. Koch. and Galium aparine L. while Boerhavia diffusa Auct Plur., Desmostachya bipinnata (L.) Stapf., Euphorbia prostrata Ait. and Rhynchosia capitata (Heyne ex Roth) D.C. complete their life cycle in early season and shed seeds. Episodic seedling emergence from a persistent seed bank of propagules is a life history characteristic that may confer reproductive advantage in unpredictable habitats so as to maximize the chance of seeding adult plant (FAO, 1994).
Some weed species were restricted only to a particular area e.g Centauria iberica Trev. ex Spreng. and Pentanema vestitum (Wall. ex DC) Ling. These were recorded only from tehsil Choa Saiden Shah. This shows an association between weeds and specific site properties. Andersson and Milberg (1998) reported that the greatest difference was found between sites and weed flora.
Plenty of motives affect the weed flora of a region. Establishment of a particular weed flora at a particular place is influenced by many factors. Amongst them are climate (Grime, 1979; Werger 1979), light, rainfall and moisture (Aldrich, 1984; Wilson, 1985; Milijic, 1987; Robert et al., 1988), temperature (Williams and Groves, 1980), nutrients (Jager and Ponso, 1979; Daniels, 1986; McGraw and Chapin, 1989; Nadeau and William, 1990), soil texture, soil moisture and soil fertility (Naeem et al., 1994; Dieleman et al., 2000b), agronomic and weed management practices, temporal weather variation and tillage system (Swanton et al., 1999; Dieleman et al., 2000a).
Besides floristic composition, biological spectrum based upon life form was also determined (Table 2). Therophytes had a major share of 62.83%, followed by hemicryptophytes (23.78%), while the least contribution was made by chameephytes (12.82%). There was one parasitic weed species i.e. O. aegyptiaca Pers. (Table 2). Life form helps in defining vegetative growth status of a species. A community dominated by therophytes would be characteristics of frequently disturbed sites (Malik and Hussain, 1990b; Smith, 1996). These can be effectively controlled by depriving them to produce seeds. Any effort for their eradication must be made before flowering. Weeds needed to be removed by the four leaf stage of the crop to prevent 10% yield loss due to weed interference (Martin et al., 2001). Annual plants invest so much energy in seed production that they do not survive after reproducing (Purves et al., 1997). Annuals are gradually replaced by perennials which are difficult to control. The environment directly influence the life forms (Packham and Harding, 1982) and there is a significant degree of uniformity in the life forms of plants within a given climatic type and prevailing structural types are related to climate (Daubenmire, 1968).
The weed species were divided into various leaf size classes. Of the recorded 78 weed species, there were 26 leptophylls, 37 nanophylls, 12 microphylls and 3 mesophylls, sharing 33.33, 47.43, 15.38 and 3.84%, respectively (Table 3). Leptophylls and nanophylls were, therefore, leading leaf size classes. Plants meet moisture stress by means of various leaf adaptations and leaf size analysis evaluates the degree of climatic similarity of the region involved. As most of the species had small leaves, such species had low food requirements and they thrive best under unfavourable conditions (Murad et al., 1995). The size attained by an expanding leaf blade is greatly influenced by the availability of warmth and moisture (Daubenmire, 1968).
The reduction in leaf size is an adaptation to water availability in arid regions as the reduction in leaf area decreases the amount of solar radiation, the plant intercepts and amount of water, the plant losses by transpiration (Smith and Smith, 1998). Majority of weeds had low frequency and density and are agronomically unimportant but as a weed they share the resources of habitat. Species which appear insignificant at the time of investigation may indicate conditions that exist at an earlier time but are not present now or they may indicate as “prediction species”, a future trend (Hanson and Churchill, 1965).
Some of the recorded species, Cynodon (Chou and Young, 1975). Dicanthium (Dirvi and Hussain, 1979), Euphorbia (Hussain, 1980), Eragrostis (Hussain et al., 1984) and Rumex (Khan, 1983) have been recognized as allelopathic weeds. C. arvense L. was considered as noxious weed in 34 states of United States (Eskelsen and Grabtree, 1995). Recently C. arvense L., Convolvulus arvensis L. and Centaurea spp. were reported among the ten most noxious weeds in United States (Skinner et al., 2000). C. album L., S. halepense L. Pers., Convolvulus arvensis L. and D. bipinnata were reported as weeds from 47, 53, 44 and 60 different countries of the world, respectively (Ashiq et al., 1996). Holm et al. (1977) placed C. dactylon Pers., C. arvensis L., S. halepense (L.) pers., Portulace oleracea L., Eleusine indica L. Gaertn. and C. album L. in 18 worst weeds of the world. Among the recorded weeds Convolvulus arvensis L., D. bipinnata L. Stapf. and C. oxycantha M.B. were also included in the ten most important weeds of Pakistan (Gafoor et al., 1987). Spiny weeds like Carthamus, Cirsium and Centaurea cause problems during harvesting while twiners like Convolvulus, Vicia, Lathyrus and Galium deform plants and reduce the light supply.
The development of weed flora involve both ecological and evolutionary processes. Species of natural flora become part of weed flora as a consequence of interspecific selection which depends upon the cropping pattern and crop competition. With continuous cropping, the duration of interspecific selection increases and a land surface develop a weed flora that characteristically reflects both time and type of cropping (FAO, 1994). Soil type and local climate conditions serve to further differentiate amongst the weed flora (Hidalgo et al., 1990).
The present findings do not quantify the occurrence, density and herbage cover of these weeds, which is helpful in determining the ecological status of these weeds. Floristic composition, therefore, must be followed by their distribution and ecology. A detailed study explaining their autecological behaviour might help in finding an effective method to eradicate them. Not a single method but an integrated weed control could be useful in controlling and reducing the weed population of the area.
REFERENCES
- Andersson, T.N. and P. Milberg, 1998. Weed flora and the relative importance of site, crop, crop rotation and nitrogen. Weed Sci., 46: 30-38.
Direct Link - Dastagir, G., Imtiaz-Ul-Haq and S. Farooq, 2000. Biological spectrum and comparison of coefficient of communities between plant communities harbouring Mai Dhani Hill, near Muzaffarabad, (AJK). Pak. J. Biol. Sci., 3: 300-302.
CrossRefDirect Link - Hussain, F., 1989. Field and Laboratory Manual of Plant Ecology. National Academy of Higher Education, Islamabad, Pakistan, Pages: 422.
Direct Link - Malik, Z.H. and F. Hussain, 1990. Weeds in the corn fields of Muzaffarabad, Azad Kashmir. Pak. J. Agric. Sci., 27: 49-55.
Direct Link - Martin, S.G., R.C. van Acker and L.F. Friesen, 2001. Critical period of weed control in spring canola. Weed Sci., 49: 326-333.
CrossRefDirect Link - Swanton, C.J., A. Sherstha, R.C. Roy, B.R. Ball-Coelno and S.Z. Knezevic, 1999. Effect of tillage systems, N and cover crops on the composition of weed flora. Weed Sci., 47: 454-461.
Direct Link