
S. M. El-Darier
Department of Botany, Faculty of Science, Alexandria University, Alexandria, Egypt
Pakistan Journal of Biological Sciences
Year: 2002 | Volume: 5 | Issue: 1 | Page No.: 6-11







ABSTRACT
The allelopathic effects of Eucalyptus leaf-litter water extract (ELWE) and crude leaf-litter powder (ECLP) were studied on growth activities, nutrient uptake and the rate of metabolite accumulation of broad bean (Vicia faba L.) and Maize (Zea mays L.). The results showed that the germination of two species (maize >broad been) was enhanced at a level of 1% ELWE and greatly reduced (broad bean > maize) at 5% level. The shoot and root lengths, total dry weight and relative growth rate (RGR) (shoot > root) were greatly reduced in broad bean than in maize. Similarly, the leaf area index (LAI) was significantly (p < 0.05) reduced in the two species. The allelopathic compounds released from ECLP significantly (p < 0.01) suppressed the uptake of N, P and K in the two examined species. Such reduction was not correlated to the concentration of nutrients in plant tissues but highly positively correlated (p < 0.01) with the reduction that occurred in dry matter production. The two species exhibit a tendency to accumulate amino acids; proline and sugars, which may be considered as an adaptive mechanism to increase the stress tolerance. The study indicates a firm conclusion for the possibility of Eucalyptus phenolic and terpenoid compounds phytotoxicity for the two examined species.
PDF Abstract XML References Citation
How to cite this article
S. M. El-Darier, 2002. Allelopathic Effects of Eucalyptus rostrata on Growth, Nutrient Uptake and Metabolite Accumulation of Vicia faba L. and Zea mays L. Pakistan Journal of Biological Sciences, 5: 6-11.
DOI: 10.3923/pjbs.2002.6.11
URL: https://scialert.net/abstract/?doi=pjbs.2002.6.11
DOI: 10.3923/pjbs.2002.6.11
URL: https://scialert.net/abstract/?doi=pjbs.2002.6.11
INTRODUCTION
The interaction of plants through chemical signals ‘allelopathy’ has many possible agricultural and ecological applications (Nelson, 1996). The interest in diversifying farm income and reducing environmental impacts on agriculture has led to the development of alley cropping systems in many districts in the Nile Valley and some irrigated areas in the north western desert of Egypt. Similar to these types of production systems, trees and or shrubs are established in hedge rows on crop land with agronomic crops, young fruit trees or pasture grasses cultivated in the alleys (Jose and Gillespie, 1998). The crops most often planted in these regions include maize (Zea mays L.), broad bean (Vicia faba L.), wheat (Triticum spp.), barley (Hordeum vulgare L.), berseem (Trifolium alexandrinum L.), potato (Solanum tuberosum L.), tomato (Lycopersicon esculentum Mill.) and other vegetables combined with trees such as Eucalyptus spp., Casuarina spp., Salix spp., Morus spp., Date palm and others. The Eucalyptus species are considered the most notorious of allelopathic trees causing understory suppression specially in dry climates and when water is scarce (rainfall<400mm) (May and Ash, 1990; Thor, 1997). Debris from the tree pruning and litter reduces agricultural yields, whereas the leaves are unpalatable and possibly allelopathic to pasture growth (Anderson, 1990). Several studies (Alexander, 1989; Kohli et al., 1988; Moore, 1997; Paulino et al., 1987) revealed that large areas of the ground surface beneath the Eucalyptus remains completely bare and ground vegetation is very limited in extent, tending to be concentrated in wetter hollows. Evidences were obtained about the progressive degradation of the soil under Eucalyptus through the increase of cation exchange capacity (CEC) and the decrease in both pH and base saturation (Alexander, 1989).
The most affected crops in terms of production in the study area are Vicia faba and Zea mays (personal communication with some indigenous farmers and visual field observations) which may be due to multiple habitat factors such as poor tillage, intensive and mixed agriculture, quality of irrigation water, past crop residues and harmful effects of neighbouring plants. The main objectives of the current work are: 1) analyzing the range of the allelopathic effects exerted by Eucalyptus rostrata on some biochemical and physiological processes of broad bean and maize and 2) to discuss the difference in tolerance of the two species to allelopathy.
MATERIALS AND METHODS
Field site: The field site of the present study includes areas of 5 feddans (460 x 460 m2) surrounded from all sides by two rows of Eucalyptus trees. Between the two rows, an irrigation canal was entrenched. This canal became as a continuous source of Eucalyptus leaf-litter extracts affecting the entire farm, besides the direct effect of Eucalyptus leaf litter falling on the ground.
Seed material: Seeds of broad bean (Vicia faba L., cv. Giza-1, family: Fabaceae) and grains of maize (Zea mays L., cv. Giza-2, family: Poaceae) were generously supplied by the Egyptian Ministry of Agriculture, El-Behira Province Authority. Laboratory experiments were conducted at the Department of Botany, Faculty of Science, Alexandria University for one growing season during 2000. Seeds and grains were thoroughly washed with distilled water and surface sterilized in 90% ethanol for 2 minutes, followed by soaking for 5 minutes in a solution of 20% commercial Clorox (5.25% sodium hypochlorite), then rinsed four times with distilled water.
Preparation of Eucalyptus leaf-litter water extract (ELWE): Ten and fifty grams of air-dried Eucalyptus leaf litter were separately soaked in one liter of double distilled water (DDW) for 12 hours at room temperature. The mixtures were filtered through Whatman No.1 filter paper to obtain 1% ‘w/v’ (Bhowmik and Doll, 1984) and 5% ‘w/v (Wardle et al., 1992) strength concentrations. The two extract levels besides the control were chosen to undergo the first experiment. The Eucalyptus leaf litter has high concentration (63.5± 16.2 mg/g) of water soluble total phenolics as determined colourimetrically (Swain and Hillis, 1959). A Randomized Complete Block design with five replicates within a block was followed.
The first experiment: Ten seeds of broad bean and ten grains of maize were arranged in 9-cm diameter petri-dishes on one disc of Whatman No. 1 filter paper under normal laboratory conditions with day temperature ranging from 20-25 oC and night temperature from 12-17 oC. Ten cm3 of each level of the Eucalyptus extract were added daily to five replicates. Double distilled water was applied to the control level. The germination rate was recorded 7 days post emergence of the radicle.
The second experiment: In this experiment Eucalyptus crude leaf-litter powder (ECLP) was used instead of leaf litter water extract applied in the first experiment. Five seeds of broad bean and five grains of maize were planted in plastic pots of 45 cm depth, 32 cm top and 20 cm bottom diameters, provided with drainage holes. Each pot contains 6 kg air-dried clay soil and treated with either of 0, 60 or 300g of ECLP applied on the surface to achieve a ratio (w/w) of 0, 1 or 5% in five replicates. After emergence, seedlings were thinned to two plants per pot to achieve planting density 25 plants/m2.
The plants were watered every two days on the average with normal tap water. The amount of water corresponding to average soil-plant evapotranspiration calculated from weight loss over a 24-hour interval. The experiment continued for forty days after which the final measurements were recorded and both species were harvested. The different organs were separated from the shoot and the leaf area was determined by a digital planimeter. Leaf area index (LAI) was calculated by dividing the total leaf area/plant on the ground leaf area specific for this plant. Organs dry weights were obtained after oven drying to constant weight at 65 oC. Relative growth rates of shoot and root (RGRs & RGRr respectively) over a period of 33 days were calculated using the formula quoted by Jose and Gillespie (1998) as follows:
![]()
The dry weights of plumule and radicle after one week from planting were considered the initial shoot and root dry weight of the studied species.
The dried shoots and roots after weighing were combined and ground in a Wiley mill and prepared for determination of N, P and K concentrations according to the methods described by Allen (1974). Total free amino acids (TFAA) and free proline (FP) were determined by the methods of Ya and Tuunekazu (1966) and Bates et al. (1973) respectively. Mono-and polysaccharides were determined according to Naguib (1963).
Data analysis: Data of growth parameters and contents of minerals and metabolites were treated with one way ANOVA test using COSTAT 2.0 Statistical Analysis Software. Comparison of the main effects was performed using Least Significant Difference (LSD) with the control.
RESULTS
Field observations: Three distinct inhibition zones were noticed within the two crop species. The first (2.5 m from tree trunk) was mostly bare from the two investigated species. The second and third inhibition zones were delimited at distances of 3.9 and 5 m from the tree row respectively. The number of broad bean individuals was 17 and 32 while that of maize was 20 and 35 in the two zones respectively. At the control site the number of individuals was 25 and 23 for broad bean and maize respectively.
Germination, growth and development: The results listed in Table 1 showed that the germination efficiency of broad bean and maize plants increased significantly (p<0.05) by about 36 and 54% respectively at 1% ELWE level relative to control plants. Contrary to this, 5% ELWE caused a reduction of about 36 and 18% from the control in broad bean and maize respectively. The shoot and root lengths at 5% Eucalyptus crude leaf-litter powder (ECLP) were lower than the control by about 64 and 67% in broad bean and about 36 and 39% in maize respectively. The individual leaf number and area and leaf area index (LAI) were greatly reduced at concentration levels of 1 and 5% ECLP.
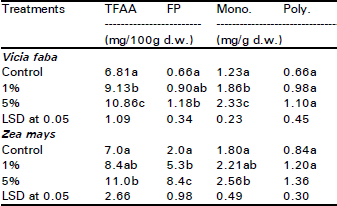
The total dry weight and shoot/root ratio were significantly reduced in broad bean (76 and 61% respectively) and maize (68 and 59% respectively) plants as affected by the two concentration levels of ECLP (Table 2). The sharing percentage of each organ to the total biomass (organ: weight ratio) differed significantly with organ and treatment (Fig. 1). At 5% treatment level, the great reduction (about 39%) for the two species occurred in leaves. The reduction in stem was greater in maize (about 38%) than in broad bean (about 9%). On contrary, the root significantly increased with the increase in treatment concentration. The relative growth rate (RGR) (g/day) was reduced in the two species (broad bean > maize) as influenced by the application of 1 and 5% ECLP (Fig. 2). At 5% level, the growth of shoot was more severely inhibited in the two species (82.5% for broad bean and 71.8 for maize) compared with root.
Nutrient uptake and metabolite accumulation: The data listed in Table 3 shows that in broad bean plants, 1 and 5% treatments of ECLP enhanced the accumulation of both P and K, while N not affected. In maize however, K increased, N dropped while P not affected. Nutrient uptake (mg/plant) is considered here as the total content [dry weight (g) multiplied by the concentration (mg/g)] of the element accumulated in the plant over a period of 40 days (Fig. 3). Significant inhibitory effects (p < 0.05) of 5% treatment of ECLP on the uptake of N, P and K was clearly observed in the two species. The reduction was about 74, 50 and 66% in broad bean and 76, 68 and 53% in maize respectively for the three elements (Table 4).
The influences of ECLP on total free amino acids (TFAA), free proline (FP), and mono- and polysaccharides in the two species are listed in Table 5. The highest effect was on FP in maize plant (4.2 times of the control). The accumulation of TFAA and polysaccharides was more or less equal in the two species. Monosaccharide accumulation reached a value of about 1.89 and 1.42 times higher than that of the control in broad bean and maize plants respectively.
DISCUSSION
The maintenance of surface residues is the main goal of conservation tillage. Allelochemicals produced and released by certain plants (through leaching, volatilization, exudation or decomposing of the remains) are one of the most important components of the stresses that may influence seed germination, plant growth and nutrient uptake of the associated plant species (Bhowmik and Doll, 1984; Rice, 1984; Shilling et al., 1985). The present study indicates that the seed germination stage of broad bean and maize is less sensitive to allelochemicals produced by Eucalyptus crude leaf-litter powder (ECLP) than the seedling stage.
| Table 1: | Germination percentage (7 days after planting) and some growth parameters (40 days after planting) in Vicia faba and Zea mays plants as affected by Eucalyptus rostrata extract and Eucalyptus crude leaf powder (ECLP) applied on the surface. |
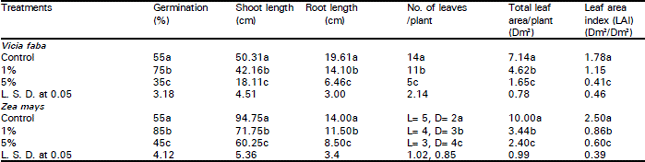 | |
| L = Foliage leaves D=Dead leaves | |
| Different letters indicate a significant difference at 0.05 level of probability as evaluated by ANOVA test. | |
| Table 2: | Influence of Eucalyptus rostrata crude leaf powder (ECLP) on the dry weight (g/plant) and shoot:root ratio of (a) Vicia faba and (b) Zea mays plants 400 days after planting |
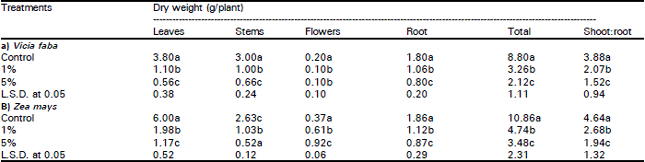 | |
| S. D. M. = Standing dead materials | |
| Different letters indicate a significant difference at 0.05 level of probability as evaluated by ANOVA test. | |
| Table 3: | Influence of Eucalyptus rostrata crude leaf powder (ECLP) (applied on surface) on the accumulation (mg/g) of N, P and K in Vicia faba and Zea mays plants 40 days after planting |
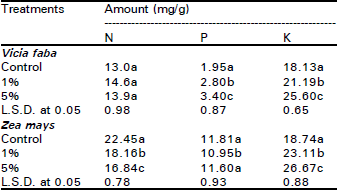 | |
| Different letters indicate a significant difference at 0.05 level of probability as evaluated by ANOVA test. | |
| Table 4: | Correlation coefficients and linear regressions between total dry weight and N, P and K uptake for Vicia faba and Zea mays plants as affected by Eucalyptus rostrata crude leaf powder (ECLP) applied on the surface |
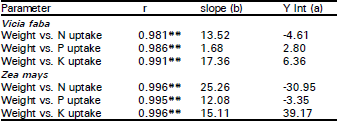 | |
| ** Values are significant at 0.01 level of probability | |
| Table 5: | Influence of Eucalyptus rostrata crude leaf powder (ECLP) on TFAA, FP, mono. and polysaccharides contents in Vicia faba and Zea mays plants, 40 days after planting |
 | |
Different letters indicate a significant difference at 0.05 level of probability as evaluated by ANOVA test. | |
| TFAA = Total free amino acids | |
| FP = Free proline | |
| Mono. = Monosaccharides | |
| Poly. = Polysaccharides | |
One percent ECLP revealed a distinct stimulatory effect on the germination of the two species (maize > broad bean). The allelopathic compounds such as fulvic, humic and cinnamic acids (polyphenols) released from Eucalyptus leaf-litter have positive effects and may be considered as promoters for the metabolic processes and seed germination if occurred at a threshold concentration (1% ECLP in the present study).
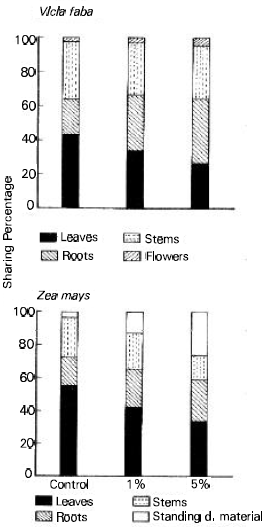 | |
| Fig. 1: | Influence of Eucalyptus rostrata crude leaf powder on the sharing percentages of the different organs to the total biomass (organ:weight ratio) of Vicia faba and Zea mays 40 days after planting. |
Moreover, such compounds alleviate the aluminum toxicity in maize, thus enhancing growth (Lambers et al., 1998). El-Darier and Youssef (2000) reported that the maximum growth rate of Lepidium sativum was attained at 25% alfalfa aqueous extract, and decreased with the increase in extract concentration. On the other hand, the nutrients released from decomposition of the ECLP at a certain concentration may play an important role in enhancing the seed germination and overcoming potential allelopathic responses of the two crop species. A similar trend was described by Indergit and Darkshimi (1994) on tomato and mustard plants and by Martin et al. (1990) on maize. The great reduction in seed germination of broad bean compared to maize at 5% ECLP may indicate the higher sensitivity of dicotyledonous species than monocotyledonous ones (Miller, 1996) and the major effect was mainly on nutrient uptake (Crawley, 1997; Lambers et al., 1998).
The two ECLP treatment levels negatively affected the growth of the two crop species in the current study. The lengths of shoot and root were greatly reduced in the two species (broad bean > maize). Similar results were obtained by Molina et al. (1991) on the effect of Eucalyptus globulus on the shoot and root lengths of Lactuca sativa.
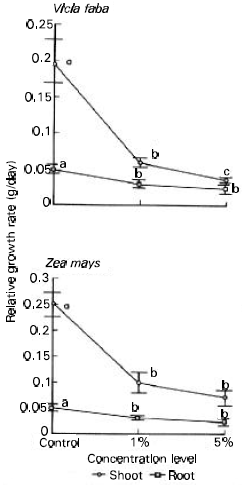 | |
| Fig. 2: | Influence of Eucalyptus rostrata crude leaf powder on the relative growth rate (g/day) of shoot (RGRs) and root (RGRr) of Vicia faba and Zea mays, 40 days after planting. Different letters indicate a significant difference at the 0.05 level of probability as evaluated by ANOVA test. |
Leaf area index (LAI) considered as one of the most important tolerance traits in maize and some leguminous plants as soybean (Lindquist and Kropff, 1996 & 1997). Moreover, Lindquist et al. (1998) reported that the growth rate of leaf area from emergence until LAI reaches 1.0 is not correlated with weed suppressive ability. In the present work the severe reduction in LAI was nearly equal in broad bean and maize plants. At 5% ECLP treatment level, the growth rate of leaf area was 0.41 and 0.60 for broad bean and maize respectively (less than 1.0) indicating a lower tolerance of the two species towards the suppressive effect of Eucalyptus. Thus increasing LAI of the crop species under allelopathic stresses by increasing population density or reducing row spacing, application of fertilizers, improving soil pH, etc. can improve the crop tolerance (Murphy et al., 1996). It is important however to bear in mind that increasing crop LAI may not be a beneficial trait in environments where water or nutrients are the most limiting (e.g. desert ecosystems), because greater leaf area may result in greater demand for water and nutrients, subsequently increasing stress during periods when resources are limiting.
The inhibition of a number of interacting physiological processes as respiration, net photosynthetic rate, transpiration and stomatal conductance is the main factor which suppressed dry matter production and growth rates (Jose and Gillespie, 1998).
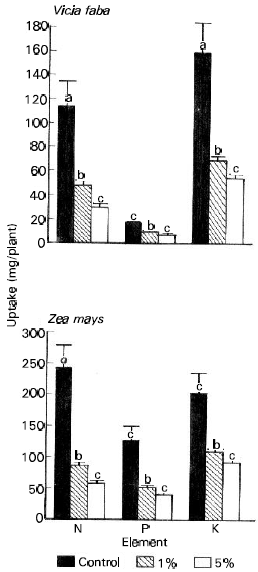 | |
| Fig. 3: | Influence of Eucalyptus rostrata crude leaf powder on N, P and K uptake (mg/plant) in Vicia faba and Zea mays 40 days after planting. Different letters indicate a significant difference at the 0.05 level of probability as evaluated by ANOVA test. |
In the present study the total dry weight (g/plant), relative growth rate (g/day) of shoots (RGRs) and root (RGRr) and organ: weight ratio (sharing percentage of the organs to the total weight) was negatively affected by ECLP treatments. The reduction in total dry weight and RGR (shoot > root) was greater in broad bean than in maize. Contrary to this, stem: weight ratio was more reduced in maize than in broad bean which may be attributed to severe reduction in shoot and root length recorded in maize. The biomass of root increased at the expense of that allocated to shoot indicating an environmental stress resulting from the release of allelochemicals. Einhellig and Leather (1988) declared that the storage of excess biomass in the root might be considered as conservative strategy to manage allelopathy.
The interruption of one plant process by the phenolic compounds released from the decomposition of Eucalyptus leaves usually affects other processes (Boes, 1986). The interference with nutrient uptake and the subsequent reduction in nutrient accumulation is one of the most effective mechanisms of phenolic compound action (Einhellig, 1987; Einhellig and Leather, 1988). This concept was confirmed in the present study where the allelopathic compounds released from ECLP significantly suppressed the uptake of N, P and K by the two studied species. Moreover, the inhibition of growth and development of the two species at higher percentage of ECLP may be due to suppressive effects on element absorption and uptake. Many polyphenols have catechol groups and hence at higher concentrations (e.g. 5% in the present study) have the ability to chelate divalent or trivalent metal ions (Crawley, 1997) and hence inhibit ion uptake (Lambers et al., 1998). Puri and Khara (1991) also obtained the same effects of Eucalyptus tereticornis on Phaseolus vulgaris. In addition, reduction in the uptake of N in maize and soybean and P in maize as affected by weed residues (Bhowmik and Doll, 1984) and also K in oat plants exposed to several phenolic acids (Harper and Balke, 1980) have been reported. Here again, the uptake of NPK was highly correlated (P < 0.01) with dry matter production which may indicate that as the nutrient uptake decreased total dry matter showed a corresponding decrease.
It is familiar that both biotic and abiotic stresses during growth often cause plants to increase production of secondary metabolites (Gershenzon, 1984) and other metabolic compounds as free acids, proline, sugars, organic solutes and others (Gorham et al., 1985). The two species of this study tend to accumulate amino acids; proline and sugars, which may be considered as an adaptive mechanism to increase, stress tolerance (El-Darier and Youssef, 1998).
In conclusion, ECLP produced inhibitory effects on broad bean and maize plants through allelopathic activity. The allelopathic effects caused by the phenolics released from Eucalyptus were mainly on crop growth due to inhibition of some interacting physiological processes such as nutrient uptake, which is highly positively correlated, to the dry matter production. The study indicated a possibility of phenolic compounds phytotoxicity for the two studied species (broad bean more sensitive than maize).
REFERENCES
- Allen, S.E., 1974. Chemical Analysis of Ecological Materials. Blackwell Scientific Publications, London, UK., Pages: 565.
Direct Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Bhowmik, P.C. and J.D. Doll, 1984. Allelopathic effects of annual weed residues on growth and nutrient uptake of corn and soybean. Agron. J., 76: 383-388.
Direct Link - Einhellig, F.A. and G.R. Leather, 1988. Potentials for exploiting allelopathy to enhance crop production. J. Chem. Ecol., 14: 1829-1844.
CrossRefDirect Link - Gorham, J., R.G.W. Jones and E. McDonnell, 1985. Some mechanisms of salt tolerance in crop plants. Plant Soil, 89: 15-40.
CrossRefDirect Link - Murphy, S.D., Y. Takubu, S.F. Weise and C.J. Swanton, 1996. Effect of planting patterns and inter-row cultivation on competition between corn (Zea mays) and late emerging weeds. Weed Sci., 44: 865-870.
Direct Link - Puri, S. and A. Khara, 1991. Allelopathic effects of Eucalyptus tereticornis on Phaseolus vulgaris seedlings. Int. Tree Crops J., 6: 287-293.
CrossRefDirect Link - Swain, T. and W.E. Hillis, 1959. The phenolic constituents of Prunus domestica. I. The quantitative analysis of phenolic constituents. J. Sci. Food Agric., 10: 63-68.
CrossRefDirect Link - Lindquist, J.L. and M.J. Kropff, 1997. Improving rice Tolerance to Barnyard Grass through Early Crop Vigour Simulation with INTERCOM. In: Systems Approaches for Sustainable Agricultural Development Applications of Systems Approaches at the Field Level, Kropff. M.J. et al., (Eds.). Kluwer Academy, Netherlands, pp: 53-62.
- Shilling, D.G., R.A. Liebl and A.D. Worsham, 1985. Rye (Secale cereale L.) and Wheat (Triticum aestivum L.) Mulch the Suppression of Certain Broadleaf Weeds and the Isolation and Identification of Phytotoxins. In: The Chemistry of AllelopathyBiochemical Interactions Among Plants. Thompson, A.C. (Ed.). ACS, Washington, DC., pp: 243-271.