
A. M. Shaheen
Department of Botany, Aswan Faculty of Science, South Valley University, Aswan, 81258, Egypt
Pakistan Journal of Biological Sciences
Year: 2002 | Volume: 5 | Issue: 11 | Page No.: 1202-1206







ABSTRACT
This study was undertaken to determine the morphological variation present within and among Ricinus communis in Egypt. The flowers, leaves, fruits, seeds and pollens of R. communis from different phytogeographical regions were examined with both scanning electron and light microscopy. Twenty two morphological characters were defined. Based on leaf, capsule, seed and pollen features such as pattern of leaf epidermal wax, stomata pattern, capsule and seed ornamentation, seed shape, seed size, seed colour, exine sculpture, aperture type, pollen shape and composition, two morphological patterns were recognized. Leaf, capsule, seed and pollen of each pattern were described, illustrated and compared. Examination of the morphological variation has shown that within R. communis two formae or varieties may be distinguished.
PDF Abstract XML References Citation
How to cite this article
A. M. Shaheen, 2002. Morphological Variation within Ricinus communis L. in Egypt: Fruit, Leaf, Seed and Pollen. Pakistan Journal of Biological Sciences, 5: 1202-1206.
DOI: 10.3923/pjbs.2002.1202.1206
URL: https://scialert.net/abstract/?doi=pjbs.2002.1202.1206
DOI: 10.3923/pjbs.2002.1202.1206
URL: https://scialert.net/abstract/?doi=pjbs.2002.1202.1206
INTRODUCTION
Traditionally fruit, seed, pollen and leaf characters have been used in delimiting subfamilies and tribes within the families of the flowering plants, i.e. in Rutaceae (Bentham, 1863; Engler, 1931; 1964). Recent scanning electron microscopic studies have provided detailed information on the surface patterns of small fruits, seeds, pollen and leaves (Heywood, 1969; 1971) and have provided a firm base for the separation of closely related species in some genera, e.g. Acacia (Shaheen, 1995). Cialdella (1984) recognized four varieties within Acacia caven, three of them were recognized by pod size and shape.
It is generally recognized that intra-specific variation is multidimensional, and that the unidimensional classification common in taxonomy is unlikely to deal satisfactory with the classifications at the intra-specific level (Heywood, 1963). The study by Lewis (1969) and Wilkins and Lewis (1964) of leaf shape in Geranium is an excellent example of how intra-specific morphological variation can be treated in an ecologically different manner. However, the study by Ayyad and Hilmy (1973) showed that the morphological variation in plants of habitats with low and high moisture availability was too wide to provide reliable criteria for the definition of taxa on an intra-specific levels.
It can also be then argued that the description of the variation at the intra specific level is essential for proper taxonomic treatment of the variation at the species level.
R. communis plants exhibit much variation regarding the size and ornamentation of the regma (fruit) and colour and patterning of the seeds, fruits, leaves and pollens. On the basis of which nearly 20 varieties and some 20 formae have been recognized (Meikle, 1985).
Ricinus L. is a monotypic genus probably native to north east tropical Africa, but now widely cultivated throughout the tropics, subtropics and warm temperate regions and often becomes naturalized presumably in the Sudanian and E. African regions (Meikle, 1985). It is considered a weed especially in the Mediterranean region, notably in Greece, Egypt, Italy and Morocco. It is subspontaneous in the hotter parts of Palestine, in the Dead sea area (Arnon River Delta) it occurs in dense groves; also common wadies, at roadsides, waste land, along the dried-up streams, abandoned gardens and in waste places (Zohary, 1987). In Egypt, it is mainly localized in the irrigated lands of the Nile valley and Delta with a few infestations in Egyptian desert (Wadi Allagi, Wadi Defiat, Egate, Gabel Elba, Sinai).
Ricinus L. is an important industrial oil plant cultivated since ancient times in tropical and subtropical countries. Up to 70% of the seed is used in the chemical industry and as an aviation lubricant; in medicine it is known as a purgative (castor oil), ricinine, a toxic alkaloid, is one of the constituents of the seed (Zohary, 1987).
This study presents an analysis of the variation in morphological features (seeds, fruits, leaves and pollens) of Ricinus communis in Egypt, it is an attempt to test the validity of taking these features as a basis of recognizing varieties or formae within the species. The aim of the study is firstly to describe in detail the seed, fruit, leaf, pollen patterns found in Ricinus in different phytogeographical regions in Egypt, and also to elucidate intra-specific variability in different phytogeographic regions of Egypt and thirdly, to assess the usefulness of the fruit, seed and pollen data for taxonomic purposes.
MATERIALS AND METHODS
Plant collections: Mature fruits, seeds, leaves and flowers (measurements were taken on samples of five mature specimens at each locality) of R. communis were collected from different
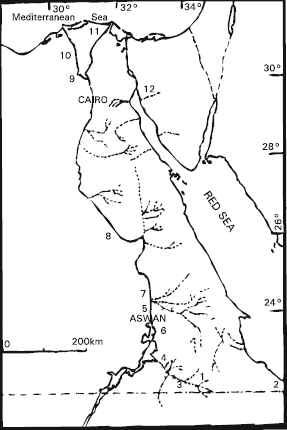 | |
| Fig. 1: | Map of the study area indicating the locations from which specimens of R. communis where collected; 1: Egate, 2: Gabel Elba, 3: Wadi Defeit, 4: Wadi Allaqi, 5: University campus, 6: El-Shalal, 7: El-Kubbania, 8: Assuit, 9: Barrages, 10: El-Mounfia, 11: New Demitta and North Sinai |
phytogeographic regions in Egypt (Fig. 1). Considering climatic, geomorphologic and phytogeographic affinities, eight more or less distinct phytogeographic (also ecological) territories are distinguished; among which, two (the desert and Nile land) have subdivisions (El-Hadidi, 1980). The fresh material was obtained from Aswan (El-Shalal and Kubbania), Eastern desert (Egate, Wadi Allagi, Wadi Defeit, Gabal Elba), Assuit, Barrages, El-Mounfia, North Sinai and New Demitta. These materials were collected during January 1999 to January 2000. Seeds from Egate were cultivated in the university campus (under arid conditions) and when the plants reached mature stage, their fruits, seeds, leaves and pollens, were also examined. All collections are stored in Aswan herbarium (ASW).
Morphological characters: Morphological data, of flower, leaf, fruit and seed characters were recorded with stero light microscope (Aswan herbarium). Scanning electron microscope (SEM) observations were made with a HIIACHI S-405A, after critical-point drying and sputter coating with gold-palladium (Glasgow university). Non-acetolyzed pollens, fresh leaves, fruits and seeds of all samples were examined. The samples were prepared as follows: cleaned stubs were first labeled and small squares of adhesive material were applied. A section of clean cover slip was placed on each stub too give smooth background, and fresh dry pollen was dusted on, fresh fruit, seed and leaf put on, and then sputter coated with gold.
The terminology used for leaf, fruit, seed and pollen description followed Erdtman (1966), Reitsma (1970) and Powell and Armstrong (1980).
Method of testing for mucilage: A standard test was devised for mucilage production and seeds of the Ricinus species were tested for this character. Seeds were soaked in tap water on filter paper in a petri dish at room temperature for 24 hours and subsequently examined either by eye or under the LM for the production of mucilage, which is usually easy to detect. The number of seeds tested in each sample varied from five to ten.
RESULTS AND DISCUSSION
Two morphological patterns (based mainly on capsule, seed, leaf and pollen characters) can be described from the 60 samples of the plant available for study; (Fig. 2) (Table 1).
The first one is represented by a small capsules with very sparse prickles and crested ridge with fine crystals wax (on the surface) and with stomata (Fig. 2 A and B). Seeds were gray with a smooth surface (Fig. 2C). The leaf is decorated with fine crystals and has a roughened surface due to irregular pattern wax; stomata at the same level with the other epidermal cells, some of them paracytic and the others surrounded by indistinguishable cells (Fig. 2, D and E). Pollen composition was monad, diad, tetrad and T-shaped; globose pollen shape, trilet aperture, perforate exine sculpture (Fig. 2 F&G); This pattern was the characteristic of the samples collected from the eastern desert (Egate, Gebel Elba, Defeit, and Allagi) and plants cultivated in University campus (Egate population); sites 1-5 respectively (Table 1).
The second pattern is represented by bigger capsules than in the first pattern, with dense prickles and prominent broad ridges with roughened wax and without stomata (Fig. 2 A’&B’).
| Table 1: | Morphological data of Ricinus communis species in different sites of Egypt |
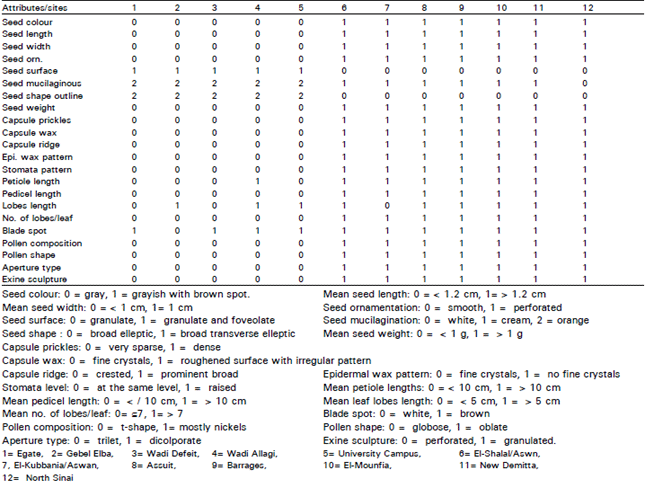 | |
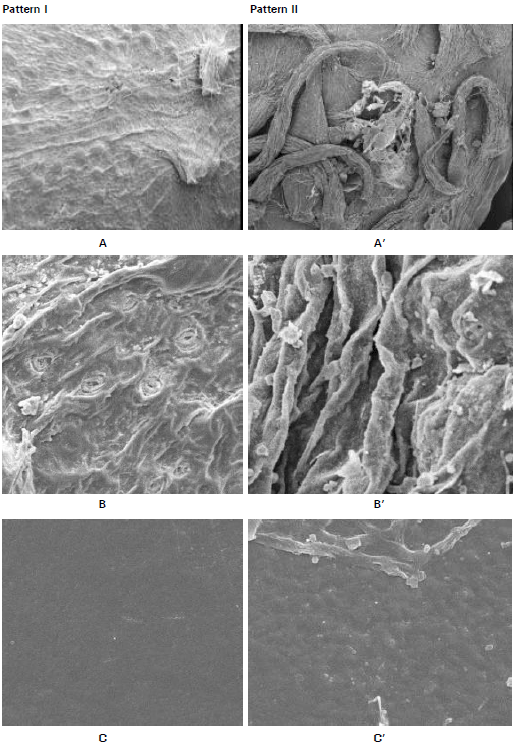 | |
| Fig. 2: | Morphological characters of Ricinus communis in different phytogeographical regions of Egypt Pattern I-II A)SEM of the capsule, showing the sparse prickles and crested ridges with fine crystals wax (location; G.E, x100). B)SEM of the capsule, showing the presence of stomata (location; G.E, x200). C)SEM of the seed, showing the smooth surface (location; G.E, x500). A’)SEM of the capsule, showing the dense prickles and broad ridges with roughened crystals wax (location; Sinai, x100). B’)SEM of the capsule, showing the absence of stomata (location; Sinai, x200). C’)SEM of the seed, showing the perforated surface (location; El-Shalal, x500). |
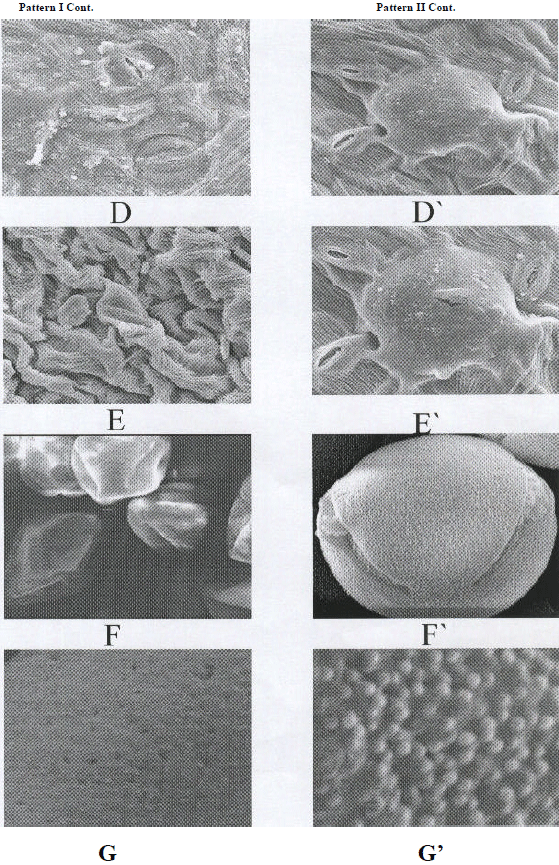 |
| Pattern II-II cont.: D&E) SEM of the leaf showing the decoration and the stomata pattern (location; G.E, x100 and 500). F&G) SEM of the pollen, showing composition and exine sculpture (location; G.E, x100 and x700). D’ &E’)SEM of the leaf showing the decoration and the stomata pattern (location; El-Shalal, x100 and 500). F’&G’)SEM of the pollen, showing composition and exine sculpture (location; Sinai, x100 and x700). |
Seeds are grayish with brown spots, and have a perforated surface (Fig. 2 C’). The leaf has roughened surface of irregular pattern wax; stomata raised above the other epidermal cells and all of them are paracytic (Fig. 2 D’&E’). Pollen composition was monad, diad, tetrad and necklace shape, oblate; dicolporate aperture and granulated exine sculpture (Fig. 2; F’ and G’). This pattern was the characteristic of the samples collected from Aswan, Assuit, El-Mounfia, Barrages, New Demitta and N. Sinai; sites 6-12 respectively (Table 1).
Morphological features (especially those of capsule, seed and pollen) in this study were obviously important for the formal recognition of intra specific taxa of R. communis and indeed there are morphological differences occur among the populations from different localities. These results agreed with those obtained by it is reported that, the electrophoretic protein patterns in roots and leaves of Ricinus communis of Egypt showed several variations (such as appearance of some newly synthesized polypeptides and disappearance of some others) between desert and riverine populations and this indicated presence of some genetic differences between them. Her study also showed several differences in the accumulation of potassium, calcium, magnesium, soluble carbohydrate, soluble protein, proline and amino acids contents in the two populations. The similar results were obtained by Meikle (1985), he found that R. communis plants exhibit much variation as regard to the size and ornamentation of the regma and the size, colour and the patterning of the seeds, fruits, leaves and pollens. On the base of which nearly 20 varieties and some 20 formae have been recognized. A near figures were obtained by Shaheen (1995) who reported that there are evidences that the populations of Acacia ehrenbergiana near the Nile and in the western desert are distinct in several characters of leaf and trunk from those in the South-Estern desert and near the Red Sea. Also there is a clear evidence of differentiation within Acacia tortilis between the South-Estern desert population and the Red Sea coastal populations, and of further divergence in individual characters within each of these two geographic zones (Shaheen, 1995). He also reported that, there is evidence of marked differentiation of the populations of Acacia nilotica correlated with latitude. However, at the specific level, fruit and seed parameters seem more conservative within populations than leaf pubescence, size of leaflets, capitulum diameter (Aronson, 1992). There is clear evidence for morphological differentiation within R. communis between the South-Eastern Desert populations and those of riverine, Deltatic and Sinai populations. The meaningful differences in morphology lead to the suggestion that R. communis may have two formae or varieties in Egypt. The morphological data should be tested against new data such as cytological, genetic, chemical data and also from outside Egypt.