
Toshiyuki Matsui
Department of Bioresource Production, Faculty of Agriculture, Kagawa University, Miki-Cho, Kagawa, 761-0795 Japan
Yousuke Hatase
Department of Bioresource Production, Faculty of Agriculture, Kagawa University, Miki-Cho, Kagawa, 761-0795 Japan
Kengo Ohobayashi
Laboratory of Biotechnology, Nagasaki Agricultural and Forestry Experiment Station, Kaizu-Cho, lsahaya, Nagasaki 854-0063 Japan
Pakistan Journal of Biological Sciences
Year: 2001 | Volume: 4 | Issue: 2 | Page No.: 228-232







ABSTRACT
We cloned and sequenced 1-aminocyclopropane-1-carboxylate oxidase (ACC oxidase, pBA-ACO) from a wounding related moso bamboo (Phyllostachys edulis) shoot using reverse transcription and polymerase chain reaction (RT-PCR) and rapid amplification of cDNA ends-PCR (RACE-PCR). The pBA-ACO was highly homologus ACC oxidase genes from submerged deepwater rice, followed by rice and banana, all of which were the wound-inducing genes in monocotyledon plants. In Northern analysis, expression of pBA-ACO mRNA was enhanced in wounding tissue until 16 hr and coincided with the peak of ACC oxidase activity.
PDF Abstract XML References Citation
How to cite this article
Toshiyuki Matsui, Yousuke Hatase and Kengo Ohobayashi, 2001. A Wound-induced ACC Oxidase Gene of Moso Bamboo Shoot. Pakistan Journal of Biological Sciences, 4: 228-232.
DOI: 10.3923/pjbs.2001.228.232
URL: https://scialert.net/abstract/?doi=pjbs.2001.228.232
DOI: 10.3923/pjbs.2001.228.232
URL: https://scialert.net/abstract/?doi=pjbs.2001.228.232
INTRODUCTION
The plant hormone ethylene, plays an important role in regulating of various physiological responses during plant growth and development. These include fruit ripening, flower senescence and abscission in response to external stresses. The biosynthetic pathway of ethylene in higher plants was well characterized and ACC synthase (EC 4.4.1.14) and ACC oxidase (EC 1.4.3) were identified. The ACC synthase is generally regarded as the rate-limiting step in the ethylene biosynthesis pathway. The oxidation of ACC to ethylene is catalyzed by ACC oxidase (called the ethylene-forming enzyme) and the role of ACC oxidase is not a major control point of ethylene biosynthesis (Yang and Hoffman, 1984). The first ACC oxidase cDNA pTOM 13 was identified in transgenic tomato (Hamilton et al., 1990) and in the expression of yeast (Hamilton et al., 1991). Subsequently, homologous sequences were identified in ripening avocado (McGarvey et al., 1990) flower senescence and elicitor treatment (Felix et al., 1991).
More recently, ACC oxidases have been isolated from ripening fruits (Ross et al., 1992; MacDiarmid and Gardner, 1993). senescing flowers (Wang and Woodson, 1992) and vegetable tissues (Peck et al., 1993; Shiomi et al., 1998; Kim and Yang, 1994). Moso bamboo is usually harvested and it is very perishable after harvest. If the ACC oxidase and ACC synthase genes of the bamboo shoot will be completely elucidated like in tomato, longer shelf life of bamboo shoot may be produced by using antisense RNA technology.
In this study, we report the cloning and characterization of the ACC oxidase gene from wounded bamboo shoot. The gene structures and the deduced peptide sequences will be analyzed and to clarify the phylogenic relationship among several ACC oxidases in other plant species.
Materials and Methods
Plant materials: Moso bamboo shoots were harvested a farmer's field in Kagawa, Japan in April 1999. The shoots were immediately brought to the laboratory, cut into 3 mm in thickness after peeling off the bracts and incubated at 25°C for 0, 8, 16 and 24 hr under humid and dark condition. After the treatment, samples were immediately stored at -80°C until needed for RNA extraction.
Enzyme activity was assayed at 30°C in 2 ml reaction mixtures containing 1.7 ml extraction buffer (without PVPP), 50 μM FeSO4, 1 mM ACC and 0.2 ml crude extract. Ethylene produced in the head space of capped tubes after 1 hr incubation was determined by gas chromatography (GC).
RNA extraction, isolation and amplification of poly (A)+ RNA: Total RNA was extracted by the ISOGEN method (Nippon gene, Osaka, Japan) according to the manufacture's protocol. Poly (A) * RNA was prepared from the total RNA by Oligo (dTI-celulose chromatography (BIO- RAD, Tokyo, Japan) as described by Watanabe (1989).
To isolate ACC oxidase fragments, degenerate oligonucleotide primers homologous to conserved regions of ACC oxidase were synthesized. The degenerate oligonuclotide primers for PCR were synthesized to two amino acid domains conserved in various ACC oxidase genes, ACENWGF for the sense primer (5'-GCNTGYGARAAYTGGGGNTT-3') and KFQAKEP for the antisense primer (5'-GGYTCYTTNGCYTGRAAYTT-31, respectively. The first strand cDNAs, synthesized from 2 g of poly (A)* RNA isolated from wound-treated fruits, were performed with a SuperScriptTM Preamplification System for First Strand cDNA (BRL, Tokyo, Japan) containing RT. In order to produce automatically cohesive end after a digestion of uracil, uracil primers containing the degenerate oligonuclotide primers mentioned above were synthesized. One is 5'-CUACUACUA-ACENWGF for the sense primer and the other is 5'-CUACUACUA-KFQAKEP for the antisense primer. The uracil primers were amplified by PCR. The parameters for PCR were 50 cycles of heating at 94°C for 30 s, at 57°C for 30 s and at 72°C for 30 s. The PCR products were then digested with uracil DNA glycosilase and cloned into the pAMP1 vector (BRL, Tokyo, Japan) by the methods of the Instruction manual. Full-length cDNA encoding bamboo cDNA was amplified by RACE method. All reactions were performed with MarathonTM cDNA Amplification Kit by the methods described in the Manual (Clonetech, Tokyo, Japan). The cDNA was synthesized from poly (A)÷RNA isolated from sliced bamboo shoots and it was ligated to the Marathon cDNA adaptor. The 5'cDNA fragment was amplified by PCR using bamboo shoot cDNA specific primer (5'-GAGGTTGACGACGATGGAGTGGCGC3') and the adaptor primer that was supplied in the kit as primers and the adaptor-ligated cDNA as a template. The 3' cDNA fragment was amplified by PCR using the bamboo shoot cDNA specific primer(5'- GGAGAACCTGGACTGGGAGAGCACC-3') and the adaptor primer as primers and the adaptor-ligated cDNA as a template. The parameters for PCR were 30 cycles of heating at 94°C for 30 s, at 60°C for 30 s and at 70°C for 3 min. The PCR products were cloned into the pSPORT 1 vector (BRL, Tokyo, Japan.
Sequencing of DNA: The cDNA inserts excised with Not I and Sal I from pAMP 1 vector and cloned into the pSPORT 1TM Not I-Sal I-cut (BRL, Tokyo, Japan). A series of deletion mutants were performed with Deletion Kit (Nippon gene, Osaka, Japan) by exonuclease III.
DNA sequencing was performed by the Cycle sequencing methods using GATCR-BioCycle sequencing Kit and a DNA sequencer GATC 1500 Long-Run system (GATC GmbH, Konstanz, Germany).
Sequence data analysis: Sequence analysis was performed using computer software from the GENETYX-MAC Version 7. Homology searches with the Genebank and the EMBL databases were performed using the Homology program in the software. The phylogenetic tree was also constructed with the UPGMA method in the software.
Preparation of the digoxigenin (DIG)-UTP-labeled RNA probe: The cloned RT-PCR product including the coding region of ACC oxidase gene was cleaved by Not I and Sal I from the pSPORT 1 vector that had been amplified in Escherichia coll (DH-5a) and it was purified by gel electrophoresis and recovered. Antisense DIG-labeled RNA probes were prepared using cloned RT-PCR product and the DIG RNA Labeling kit (Bohringer Mannhaim) according to the manufacture's instruction.
Northern blot hybridization: Ten dug Poly(A)+ RNA was subjected to electrophoresis on a 1.0% agarose gel that contained 0.66 M formaldehyde and transferred to Hybond N+ (Amersham) by capillary action with 20 x SSC, according to manufacture's instruction. After drying the membrane, the RNAs were fixed with UV. The membrane was preincubated in 5xSSPE, 5xDenhart's solution and 0.5% SDS at 60°C for 3 hr and then probed with the genespecificantisense DIG-labeled RNA probe at 60°C for 24 hr. The membrane was washed once with 2 x SSPE containing 0.1% SDS for 10 min at room temperature, once with 1 x SSPE plus 0.1% SDS for 15 min at 65°C and twice with 0.2 x SSPE containing 0.1% SOS for 15 min each at 65°C. After allowing sheep anti-DIG Fab fragments conjugated to alkaline phosphatase to bind to the DIG-labeled probe, the excess antibody was removed. Signals were detected by color reaction using 5-bromo-4-chloro-3-indol phosphate and nitroblue tetrazolium salt as the substrate. RNA samples were ensured by measuring UV spectra meter (UV-1200, Shimazu, Japan) and hybridizing an internal control gene, tomato action Cdna.
Results
Isolation and identification of cDNA: The cDNA clone of pBA-ACO was 1375 by in length and contained a 5'- untranslated region of 101 bp, an open reading frame of 966 by encoding 322 amino acid and 3'-unsaturated region of 303 by (Fig. 1). The alignment of deduced amino acid sequence showed that ACC Oxidase gene was highly homologous to the others isolated from other plant (Fig. 2). pBA-ACO had the highest percentage of sequence homology with submerged deep water rice (X85747), followed by rice ( AF049889) and banana (Liu et al., 1997; U80233) 90.2, 79.5 and 75.0% at nucleotide and 87.0, 74.0 and 74.7% at amino acid respectively, all of which have been to be wound-inducing genes (Table 1).
| Table 1: | Percentage of nucleic acid and amino acid identity between ACC oxidase from bamboo shoot and the other plants in databases |
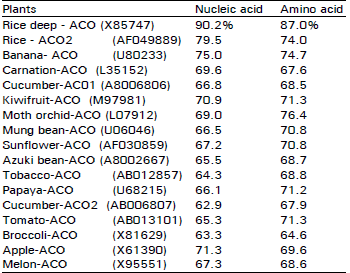 | |
| Bamboo shoot -ACO (AB044747) is calculated as 100% | |
Expression of ACC Oxidese gene: The expression of pBA-ACO was induced by wounding. Maximum pBA-ACO expression appeared 16 hr after wounding and coincided with the peak of ACC oxidase activity (Fig. 3, 4).
A phylogenetic tree was reconstructed from an alignment of the deduced amino acid sequence from the moso bamboo shoot ACC oxidase with other ACC oxidase in the database.
The pBA-ACO (accession No. AB044747) , ACO from rice submerged deepwater ( X85747) and moth orchid (L07912) clustered together strongly, with the closest relationship moso and the submerged rice. Other wounded-associated ACC oxidase are indicated in Fig. 5.
Rice and bamboo shoot belong to monocotyledon plant and other broccoli and cucumber belong to dicotyledon plant.
DISCUSSION
We have isolated a cDNA from bamboo shoot which is wounding-related. This clone, pBA-ACO, is highly homologous to a cDNA clone from rice submerged deepwater that is also associated with wounding. Tomato pTOM13 and apple pAP4 (Ross et al., 1992) are considered to be member of a small multigene family since their sequence homology were higher and Southern analysis of genomic DNA revealed multiple bands hybridising to pAP4 insert.
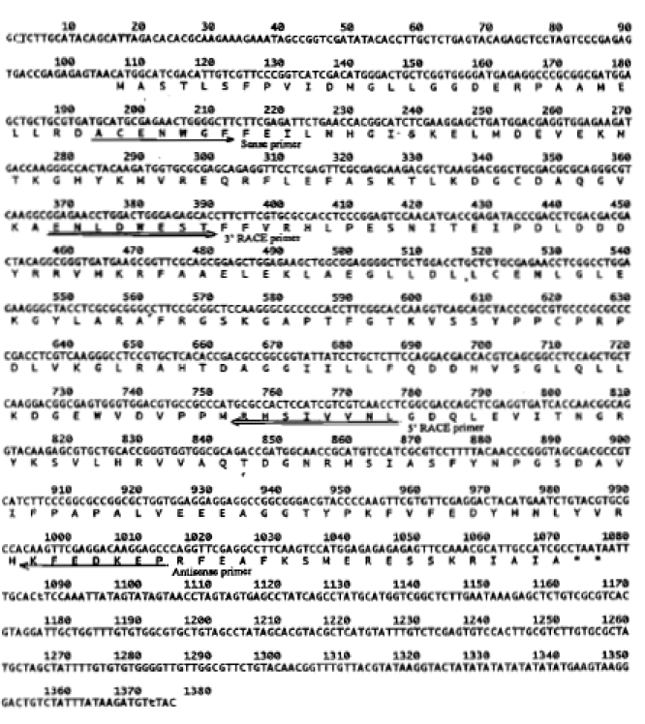 | |
| Fig. 1: | Nucleic acid sequence and drived polypeptide sequence of the pBA-ACO cDNA. The arrows indicate the positions of degenerated primers used for RT-PCR and RACE-PCR. |
The predicted polypeptide pAP4 showed 79.7% of sequence homology with pTOM13. In bamboo shoot, Northern analyis of the cDNA revealed that bands hybridising to pA134 (Data not shown) and comparison of the peptide by apple (pAP4) and bamboo shoot (pBA-ACO) genes indicated a 69.9% identity. This identity showed a higher percentage homology in comparison with different cotyledon. If the different plants belonging to monocotyledon such as bamboo shoot and rice (Table 1), the idenity showed 74.0 to 87.0% peptide sequence. This suggests that pBA-ACO may also be part of a multigene family.
Expression of pBA-ACO complementary RNA was enhanced in wounding bamboo shoot tissue. This result was consistent with that observed in apple, tomato and cucumber (Shiomi et al., 1998). In bamboo shoot, the time lag between wounding treatments and gene expression appeared to be longer compared with that in cucumber.
Phylogenetic analysis of ACO polypeptide showed that there were two major branches in phylogenetic tree, monocotyledon and dicotyledon. These are also divided to wounding, ripening and auxin induced gene. pBA-ACO belonged to the wounding-induced monocotyledon.
 | |
| Fig. 2: | Alignment of amino acid sequences deduced from bamboo (AB044747), rice deepwater (X85747), banana (U80233), moth orchid (L07912), rice (AF049889) and apple (X61390) |
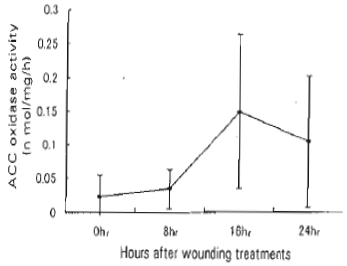 | |
| Fig. 3: | Changes in ACC oxidase activity after wounding treatment of bamboo shoot. Each point represents the mean of 3 replications. The bar shows the standard error |
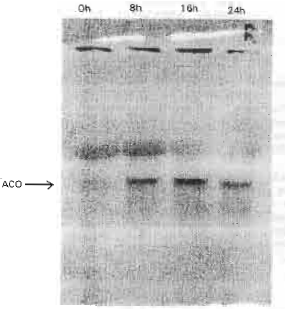 | |
| Fig. 4: | Northern analysis of total RNA (10 μg per lane) expressed from bamboo shoot following wounding treatments. RNA was extracted from tissue after 0, 8, 16 and 24 hr of wounding treatments |
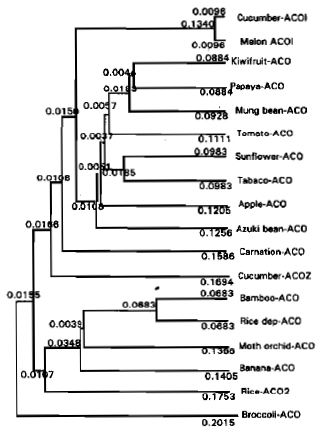 | |
| Fig. 5: | Phylogenetic analysis of ACC oxidase amino acid sequences from bamboo shoot and other ACC oxidases in the database. The phylogenetic tree was constructed from an optical alignment of proteins using the UPGMA method of GENETIX-MAX software. Accession numbers of sequences (from top to bottom of the tree): AB006806, X95551, M97961, U68215, U06046, AB0133101, AF030859, AF012857, X61390, AB002667, L35153, AB006807, A8044747, X85747, L07912, U80233, AF049889, X81629 |
*Footnote: The nucleotide sequence data reported in this paper will appear in DDBJ, EMBL and GeneBank, nucleotide sequence databases under the accession number AB044747.
ACKNOWLEDGMENTS
We acknowledge the generous gift of the pAP4 clone from Dr. Michael Lay-Yee of The Horticulture and Food Research Institute of New Zealand and that of tomato actin from Dr. Yasutaka Kubo of Okayama University. We also thank Dr. Kubo for his helpful advice.
REFERENCES
- Felix, G., D.G. Grosskopf, M. Regenass, C.W. Basse and T. Boller, 1991. Elicitor-induced ethylene biosynthesis in tomato cells: Characterization and use as a bioassay for elicitor action. Plant Physiol., 97: 19-25.
CrossRefPubMedDirect Link - Fernandez-Maculet, J.C. and S.F. Yang, 1992. Extraction and partial characterization of the ethylene-forming enzyme from apple fruit. Plant Physiol., 99: 751-754.
CrossRefDirect Link - Hamilton, G., W. Lycett and D. Grierson, 1990. Antisense gene that inhibits synthesis of the hormone ethylene in transgenic plants. Nature, 364: 284-287.
CrossRefDirect Link - Hamilton, A.J., M. Bouzayen and D. Grierson, 1991. Identification of a tomato gene for the ethylene-forming enzyme by expression in yeast. Proc. Natl. Acad Sci. USA., 88: 7434-7437.
Direct Link - Kim, W.T. and S.F. Yang, 1994. Structure and expression of cDNAs encoding 1-aminocyclopropane-1-carboxylate oxidase homologs isolated from excised mung bean hypocotyls. Planta, 194: 223-229.
CrossRefDirect Link - MacDiarmid, C.W. and R.C. Gardner, 1993. A cDNA sequence from kiwifruit homologous to 1-aminocyclopropane-1-carboxylic acid oxidase. Plant Physiol., 101: 691-692.
Direct Link - McGarvey, D.J., H. Yu and R.E. Christoffersen, 1990. Nucleotide sequence of a ripening-related cDNA from avocado fruit. Plant Mol. Biol., 15: 165-167.
CrossRefDirect Link - Peck, S.C., D.C. Olson and H. Kende, 1993. A cDNA sequence encoding 1-aminocyclopropane-1-carboxylate oxidase from pea. Plant Physiol., 101: 689-690.
Direct Link - Ross, G.S., M.L. Knighton and M. Lay-Yee, 1992. An ethylene-related cDNA from ripening apples. Plant Mol. Biol., 19: 231-238.
CrossRefDirect Link - Shiomi, S., M. Yamamoto, T. Ono, K. Kakiuchi and J. Nakamoto et al., 1998. CDNA cloning of ACC synethase and ACC Oxidase genes in cucumber fruit and their differential expression by wounding and auxin. J. Jap. Soc. Hortic. Sci., 67: 685-692.
Direct Link - Wang, H. and W.R. Woodson, 1992. Nucleotide sequence of a cDNA encoding ethylene forming enzyme from petunia corollas. Plant Physiol., 100: 535-536.
Direct Link