
Mohammad Arif
Department of Plant Pathology, Faculty of Crop Protection Sciences, N.W.F.P. Agricultural University, Peshawar, Pakistan
Sher Hassan
Department of Plant Pathology, Faculty of Crop Protection Sciences, N.W.F.P. Agricultural University, Peshawar, Pakistan
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 12 | Page No.: 2126-2130







ABSTRACT
Soybean Mosaic potyvirus (SMV) is highly prevalent in major soybean growing areas of the NWFP. Infected plants exhibited predominantly mosaic and mottling symptoms on leaves and stunted growth due to shortened petioles and internodes. Incidence of the virus ranged from mild to severe infection between 8-72% in areas surveyed. ELISA test further confirmed the incidence of the virus from 7 to 64% in major soybean growing areas of the province, during summer 1997-98. Twelve soybean mosaic virus isolates were collected from soybean crop grown in different areas on the basis of characteristic and distinguishable mosaic and mottling symptoms. Isolates, SMV-S1, S2, S3 from Swat and SMV-P1, P2, P3 from Peshawar were further selected on the basis of severity of symptoms and characterized. SMV-S1, 52, 53 produced severe mosaic on mechanically inoculated soybean (Glycine max) cv. Weber-84, leaf chlorosis and chlorotic local lesions on Phaseolus valgaris and P. lunatus and local lesions on Chenopodium amaranticolor and C. quinoa. SMV-P1, P2 and P3 isolates differed from Swat isolates by producing mild mosaic on soybean cv. Weber-84. Isolate SMV-S1 produced severe mosaic, followed by severe mottling, necrosis and occasionally death of plants Weber-84 inoculated with serially diluted sap from infected soybean plants while SMV-P1 caused mild to moderate mosaic symptoms. The dilution end points of both isolates was 11512. Mean A405,,,,, ELISA values of SMV-S1, 52 and S3 was 1.919 and of SMV-P1, P2 and P3 was 1.377, respectively, after overnight incubation of antigen with substrate solution at 4°C. Among these isolates, SMV-S1 showed maximum A405,in values (1.987) followed by SMV-P1 (1.477), respectively.
PDF Abstract XML References Citation
How to cite this article
Mohammad Arif and Sher Hassan, 2000. Occurrence and Distribution of Soybean Mosaic Potyvirus in Soybean Crop of North-west Frontier Province, Pakistan and Characterization of Prevalent Isolates. Pakistan Journal of Biological Sciences, 3: 2126-2130.
DOI: 10.3923/pjbs.2000.2126.2130
URL: https://scialert.net/abstract/?doi=pjbs.2000.2126.2130
DOI: 10.3923/pjbs.2000.2126.2130
URL: https://scialert.net/abstract/?doi=pjbs.2000.2126.2130
INTRODUCTION
Soybean mosaic virus (SMV) is one of the most prevalent viral diseases of soybean Glycine max (L.) Merrill in the world as well as in Pakistan. Yield losses due to virus infection depends upon virus strains, host genotype and time of infection. However, the virus causes 35-50% crop loss under natural infections (Ross, 1977) and as high as up to 93% in experimentally inoculated plants (Sinclair and Backmann, 1989). The virus is transmitted through seed and different aphid species in non persistent manner (Bowers and Goodman, 1979) and is also readily transmitted by sap inoculation in susceptible soybean cultivars and few other leguminous hosts (Bos, 1972). The virus produces pronounced mosaic, mottling and necrotic lesions on leaves. in some tolerant varieties inspite of infection, symptoms are riot produced due to latent infection (Lim, 1985).
Soybean mosaic virus is a serious threat to economic production of soybean in many parts of the world, particularly in Japan (Koshimizu and lizuka, 1963), Brazil (Porto and Hagedorn, 1975), United States of America (Lee and Ross, 1972; Hill et al., 1987), Korea (Cho and Goodman, 1979), Indonesia and China (Ruesink arid Irwin, 1986), Egypt (Omar et al., 1986), Argentina (Laguna et al., 1987), Thailand (Arunin, 1978; Tsuchizaki et al., 1987) and Yogoslavia (Taraku et al., 1988). SMV also occurs in other parts of the world wherever soybean is grown due to seed borne nature, where the infected seed play a pivotal role in epidemiology of the virus.
Soybean mosaic virus is transmitted in nature by insect vector belonging to the family Aphididae (Abney et al., 1976). Some 16 aphid species including Acyrthosiphon pisum, Aphis faba and Myzus persicae have been reported to transmit the virus in a non-persistent manner (Bos, 1972).Non-vector transmission is by mechanical inoculation but seed transmission is the most important source of primary infection (Bos, 1972). Up to 30% or more of the seeds of diseased plants are infected depending on cultivar and duration of infection before flowering (Bos, 1972). Plants infected after flowering do not produce infected seeds. The virus is present in seed coat and embryo and green seeds contain more virus than mature ones (Koshimizu and lizuka, 1963). Seed coat mottling is caused by virus infection but there is no correlation between mottling and the concentration of the virus in the seed (Kennedy and Cooper, 1967; Bos, 1972; Sinclair and Backmann, 1989).
Soybean mosaic virus, a species of the potyvirus genus, contain flexuous rods with modal length of about 750x15-18 nm (Galvez, 1963; Ross, 1967; Soong and Milbrath, 1980). Virus particles, ranging from 300 to 900 rim long have been also reported (Soong and Milbrath, 1980). Infectivity is positively correlated to particle size, thus the most infectious particles being over 656 nm long (Sinclair and Backmann, 1989). Nucleic acid in SMV virions have single stranded RNA, constituting 5.3% of the particle mass and having a molecular weight of 3.25 x 106d (Hill and Benner, 1980a, b).
Before initiation of these studies, very little was known about the incidence, losses, sources of resistance of SMV in Pakistan. According to preliminary studies conducted by Ali and Hassan (1993), the virus has been reported from various soybean growing regions of the N. W. F. P. However, detailed etiological and epidemiological studies have not been conducted so far. This paper reports the occurrence and distribution of SMV in major soybean growing areas of the N.W.F.P., Pakistan and identification and characterization of prevalent virus isolates in soybean crop.
MATERIALS AND METHODS
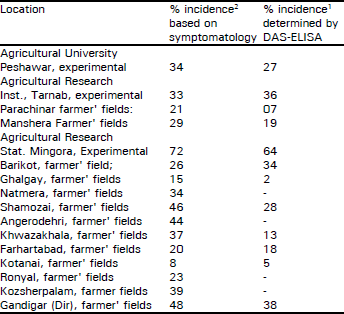
Field Survey, Virus Incidence and Distribution: Field surveys were conducted during June to September 1997-98 in major soybean growing of the NWFP in order to determine incidence and distribution of the virus (Table 1).
| Table 1: | Incidence of soybean mosaic virus in NWFP |
 | |
| 1500-600 plants were observed on each location and tested by DASELISA. 2Not tested | |
A few locations were visited even before initiation of these studies to assess the level of infection and nature of symptoms in farmer fields. Two to three plots, measuring 4×4 m2, were randomly selected in each field and three fields were selected per location and incidence of the virus was determined as the percentage of infected to total plants. Samples were collected from infected and healthy plants and tested by DAS-ELISA to determine the incidence of the virus on the basis of antigen-antibody reaction.
Virus Isolates: Collection, Preservation and Maintenance: Soybean mosaic virus infected samples were collected and on the basis of severity and variation of symptoms, 12 isolates were selected from different soybean growing areas of the NWFP (Table 2) and maintained on soybean cv. Swat-84 and Weber-84 under screen house conditions.
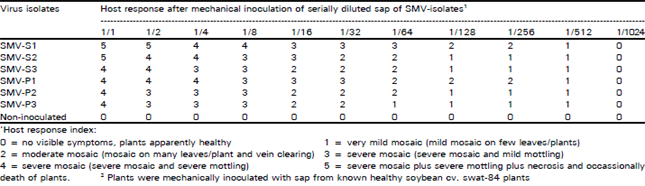
Leaf samples from infected soybean plants were collected and stored in polythene bags. Symptoms were recorded and diseased specimens were specified with respect to source of seed and variety. Virus culture was maintained on soybean cv. Weber-84 by applying standard techniques of mechanical inoculation reported by Cho and Goodman (1982). The plants were marked with date of inoculation and source of virus isolate. Control plants were inoculated with distiled water or sap from healthy plant or even with buffer. The biological purity of the isolates were confirmed by serial passage on Phaseolus vulgaris cv. Top Crop and then back to soybean cv. Weber-84. Pure SMV isolates were kept in insect proof screen house and maintained by sub-culturing the isolates on to healthy soybean plants. Seeds were collected from infected plants and kept at 4°C and used as a virus source in future. The host reaction to different SMV-isolates was evaluated by using a modified disease indexing scale as reported earlier by Ilyas et al. (1992) as: 0= no visible symptoms, plants apparently healthy; 1 = very mild mosaic (mild mosaic on few leaves/ plants); 2 = moderate mosaic (mosaic on many leaves/ plant and vein clearing); 3 = severe mosaic (severe mosaic and mild mottling); 4 = severe mosaic (severe mosaic and severe mottling); 5 = severe mosaic plus severe mottling plus necrosis and occasionally death of plants.
Plant Culture and Growth Conditions: Seed was germinated in a sterile mixture which consisted of field soil, peat, sand and farm yard manure (1:1:1:1 vlv) in 26 Cm diameter clay pots and were kept in insect free screen-house at 25-30°C.
Serological Detection of the Virus
Indirect ELISA: Indirect ELISA was performed in polystyrene micro-plate (NUNC Immunoplate II) containing 96 wells. Wells were coated with 200 μl aliquots of tissue extract from infected and healthy samples, diluted (1:5 w/v) in 0.05 M carbonate coating buffer, pH 9.6 (1.5 g Na2CO3, 2.93 g NaHCO3 and 1000 ml distiled water) and incubated for one hour at 37°C. Wells were rinsed with phosphate buffered saline (PBS), pH 7.4 containing 0.05% Tween-20. Specific antiserum (200 μl/well at 1:1000 (v/v) in Indirect ELISA buffer (IE8:0.01 M PBS, pH 7.4, 0.05% Tween-20, 2% polyvinylpyrrolidone (Sigma Chemical Co., USA) and 0.05% egg albumin (Fluke G., Switzerland). Plates were incubated in the incubator for 30 min. at 37°C. After rinsing with P85 Tween-20 as before, plates were coated with 200 μl aliquots of goat anti-rabbit immunoglobulin conjugated to alkaline phosphatase (Sigma Chemical Co., USA) diluted at 1:1000 dilution in IEB and incubated for 30 min. at 37°C. After rinsing with PBS-Tween-20, bound enzyme conjugate was detected 30-60 min. after addition of substrate buffer at 200 μl/well (p-nitrophenyl phosphate at 0.6-1 mg/ml in 0.1 M diethanolamine buffer, pH 9.8). Plates were read at absorbance (A405 nm) with Titertek Multiskan, Model MC (Flow Laboratories Inc.). Samples were considered to be positive when A405 values exceeded the mean of the virus-free samples by at least a factor of two.
DAS-ELISA: DAS ELISA was done by using PathoScreen Kit (Agdia, Elkhart, Indiana, USA) and test was performed as per instruction provided with each kit. Plant samples were collected and sap was extracted in plastic bags by rolling a marker or using pestle and mortar in extraction buffer, pH 7,4 (1000 ml of extraction buffer was prepared by dissolving sodium sulphate (anhydrous 1.3 g), PVP (MW 24-40,000 20.0 g), sodium azide (0.2 g), powdered egg (chicken) albumin (grade II 2.0 g), Tween-20 (20.0 g) in 1x PBST. 1:10 (v/v) or tissues were ground in extraction buffer at 1:10 ratio (w/v). 100 homogenate was despensed in pre-coated ELISA plates (Agdia, Elkhart, Indiana, USA). Same amount of sap from known SMV infected and known healthy soybean plants was loaded in each positive and negative control wells. ELISAplates were incubated inside a humid box for 2 h at IRT or overnight in refrigerator at 4 nc. Plates were washed with 1×PBST buffer pH 7.4 (1000 ml of 1×PBST: sodium chloride 8.0 g, sodium phosphate dibasic (anhydrous) 1.15 g, Potassium phosphate mono-basic (anhydrous) 0.2 g, potassium chloride 0.2 g, Tween-20 0.5 g was dissolved in distiled water to make the volume up to 1000 ml 4 to 8 times. After washing, 100 A.41 of enzyme conjugate was dispensed in each well of the plate and incubated in a humid box for 2 h at RT. The plates were washed 4 to 8 times with 1×PBST. 100 of OPD solution, pH 5.0 (100 ml of OPD solution was prepared by dissolving hydrogen peroxide (30%) 0.4 ml, citric acid (anhydrous) 5.1 g, sodium phosphate, dibasic (anhydrous) 7.33 g in 900 ml of distiled water and volume was adjusted to 1000 ml by adding more distiled water) per well was added and the plates were incubated 15-30 min in humid box at FIT or overnight (i.e. 16 h) at 4°C. The reaction was stopped by adding 50 μI of 3M sulfuric acid to each well.
| Table 2: | Soybean mosaic virus isolates collected from soybean growing areas of the NWFP |
 | |
1-2SMV-S1 collected from experimental fields at Agricultural Research Station, Mingora, Swat during survey conducted and mechanically transferred on Soybean cv. Swat-84 and Weber-84 at screen house conditions. 3SMV-3 isolated from infected soybean plant cv. Weber-84 from Shamozai, Swat. 4-5SMV-P1 and SMV-P2 were isolated from infected soybean cv. Weber-84, NWFP Agricultural University, Experimental at Malakandhre and mechanically transferred on soybean cv. Swat-84 in screen house. 6SMV-P3 isolated from infected soybean plant cv. Weber-84, from Agricultural Research Institute, Tarnab, Peshawar and maintained on soybean cv. Swat-84 in screen house. | |
| Table 3: | Biological characterization of soybean mosaic virus isolates |
 | |
| Table 4: | Virulence of serially diluted sap of six soybean mosaic virus isolates on Weber-84 |
 | |
| Table 5: | Serological characterization of soybean mosaic virus isolates |
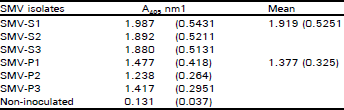 | |
| 1Mean of three replicated wells. A405 after overnight (i.e 16 h) at 4°C Ireadings after only 1 h at noon; temperature in parentheses). 2 Non-inoculated, virus-free, soybean cv. Weber-84 | |
The reaction was assessed visually or measured at 490 nm with Titertek Multiskan, Model MC Flow Laboratories Inc.). The samples were considered to be positive when the A490 values exceeded the mean of the virus-free samples by at least a factor of two.
RESULTS
incidence and Distribution of Soybean Mosaic Virus in NWFP: Soybean mosaic virus occurred widely in major soybean growing areas of the NWFP, Infected plants exhibited predominantly mosaic and mottling symptoms on leaves and stunted growth due to shortened petioles and internodes (Fig. 1) Virus infection ranged from mild to severe infection with the incidence between 8-72% (average of 40%) in areas surveyed (Table 1). ELISA test further confirmed the incidence of the virus which was 7-64% (average of 35.5%) in major soybean growing areas of the province, during summer 1997-98 (Table 1). Highest incidence of the virus (72%) was recorded in experimental plots of Agricultural Research Station, Mingora, on the basis of characteristic symptoms and ELISA (54%) whereas lowest (8% and 5%), respectively in a farmer field near Kotanai, Distt. Mardan (Table 1).
Collection, identification and biological characterization of soybean mosaic virus isolates: Twelve soybean isolates were collected from soybean crop grown in different areas, which caused characteristic mosaic and mottling symptoms. Out of these, six isolates, three each from Swat (SMV-S1, S2, S3) and Peshawar areas (SMV-P1, P2, P3), were selected on the basis of severity of symptoms for further studies and characterization (Table 2). SMV-S1, S2, S3 produced severe mosaic on mechanically inoculated soybean cv Swat-84 and Weber-84, leaf chlorosis and chlorotic local lesion on Phaseolus valgaris and P. lunatus and local lesions on Chenopodium amaranticolor and C. quinoa. SMV-P1, P2 and P3 isolates exhibited mild mosaic reaction on soybean cv. Weber-84 and similar reaction was recorded in P. vulgaris and Chenopodium species as reported earlier (Table 3).
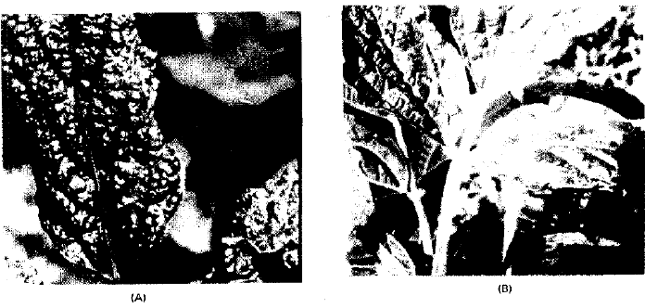 | |
| Fig. 1: | Soybean germplasm infectd with soybean mosaic potyvirus. Soybean leaves infected with SMV -S1 isolate showing mosaic and severe mottling (puckering symptoms (A) and soybean leave infected with SMV-P1 isolate showing mosaic and chlorosis symptom (B) |
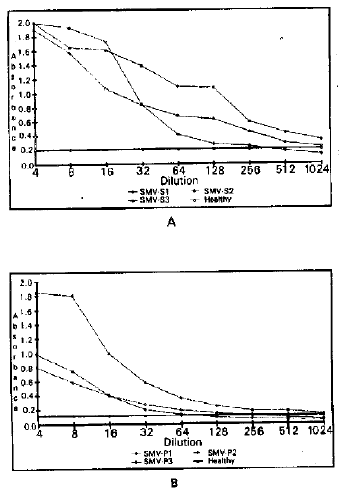 | |
| Fig. 2: | Absorbance values (A405nm) obtained in ELISA of serial dilutions of extracts from plant infected with SMV-isolates S1, S2, S3 (A) and SMV-isolates P1, P2, P3 (B). A 405nm of SMV-S1 (a) and SMV-P1 (b) are highest among other isolates. |
Soybean mosaic virus isolate S1 (SMV-S1) produced severe mosaic, followed by severe mottling, necrosis and occasionally death of plants of soybean cv. Weber-84 inoculated with serially diluted sap of systemically infected soybean plants. SMV-P1 isolate had a moderate host response. The dilution end points of both isolates was 1/512 (Table 4).
Serological characterization of soybean mosaic virus isolates: Virus isolates from infected sap of three weeks mechanically inoculated soybean cv. Weber-84 plants were also tested by DAS-ELISA. The mean A405nm absorption values of SMV-S1, 52 and 53 was 1.919 and SMV-P1, P2 and P3 was 1.377, respectively, after 16h incubation of antigen with substrate solution at 4°C. Among these isolates, SMV-S1 showed maximum A405nm values (1987) followed by SMV-P1 (1.477), respectively (Table 5). In tests on serial dilutions of infected plant extracts of SMV isolates such as SMV-S1, 52 and S3 were detected up to a dilution of 1/512, 1/64 and 1/256, respectively, whereas SMV-P1, P2 and P3 isolates were detected in serial dilutions up to 1/28, 1/16 and 1/32, respectively. SMV-51 could be detected in highest serial dilution of infected plant extract followed by SMV-P1 (Fig. 2).
DISCUSSION
Soybean mosaic virus is one of the most economically destructive viruses in soybean and its transmission through seed plays a vital role in disease epidemiology and virus spread. Present studies revealed that the virus is widely prevalent in all soybean growing areas of the NWFP. However, per cent infection and severity of the disease depends upon time of infection, virus strain, prevalent climatic conditions and host genotype (Goodman et al., 1979; Irwin and Goodman, 1981).
Our studies further revealed that the virus produced severe symptoms in soybean crop in Swat areas, particularly in experimental fields of Agricultural Research Station (North), Mingora, than in Peshawar. Possible reason of this variation is the temperature. Temperature in Swat remained between 15-18°C during growing season which is conducive for virus multiplication and development of symptoms whereas symptoms remained masked at higher temperature (>25°C) (Sinclair and Backmann, 1989). Mean temperature at Peshawar was above 30°C during the growing season.
Incidence of the virus was also high in experimental fields of ARS, Mingora, Swat. This may probably be due to sowing of virus infected seeds, as the virus is seed-borne (Porto and Hagedorn, 1975; Zhang et al., 1986) and thus high incidence of the virus in infected plants is more logical. Similar explanation have been given for SMV in soybean (Goodman and Oard, 1980) and in other crops by seed transmitted viruses. High incidence of cucumber mosaic virus in Phaseolus vulgaris germplasm (Davis et al., 1981), pea-seed borne virus in lentil germplasm (Hampton, 1982), urd bean leaf crinckle virus in mung bean (Vigna radiate) germplasm (Beniwal et al., 1980) and bean common mosaic virus in urd bean (Vigna mungo) germplasm (Agarwal et al., 1979) has been reported. Two SMV-isolates (SMV-S1 and SMV-P1), we selected on the basis of severity of symptoms and some serological properties may not cover whole pathogenic variation of SMV. The possibility of pathogenic variation of SMV still exist in this area and other part of Pakistan. More work is needed to exploit this pathogenic variation on the basis of coat protein and nucleic acid sequencing, analysis and comparison with other SMV isolates.
ACKNOWLEDGMENTS
The financial support of Pakistan Science Foundation is gratefully acknowledged.
REFERENCES
- Abney, T.S., J.O. Sillings, T.L. Richards and D.B. Broersma, 1976. Aphids and other insects as vectors of soybean mosaic virus. J. Econ. Entomol., 69: 254-265.
Direct Link - Cho, E.K. and R.M. Goodman, 1982. Evaluation of resistance in soybeans to soybean mosaic virus strains. Crop Sci., 22: 1133-1136.
Direct Link - Davis, R.F., Z. Weber, H. Pospieszny, M. Silbernagel and R.O. Hampton, 1981. Seedborne cucumber mosaic virus in selected Phaseolus vulgaris germ plasm and breeding lines in Idaho Washington and Oregon. Plant Dis., 65: 492-494.
Direct Link - Goodman, R.M., G.R. Bowers Jr. and E.H. Paschal, 1979. Identification of soybean germplasm lines and cultivars with low incidence of soybean mosaic virus transmission through seed. Crop Sci., 19: 264-267.
Direct Link - Hill, J.H., T.B. Bailey, H.I. Benner, H. Tachibana and D.P. Durand, 1987. Soybean mosaic virus: Effects of primary disease incidence on yield and seed quality. Plant Dis., 71: 237-239.
Direct Link - Omar, R.A., F.F. Mehiar, E.A. Zayed and A.A. Deif, 1985. Biological studies on some seed-borne viruses and their effect on vegetative growth and yield component of the host plants. Acta Phytopathol., 20: 59-78.
Direct Link - Ross, J.P., 1977. Effect of aphid-transmitted soybean mosaic virus on yields of closely related resistant and susceptible soybean lines. Crop Sci., 17: 869-872.
Direct Link