
Asghar Ali
Pulses Programme, National Agricultural Research Centre, Park Road, Islamabad, Pakistan
C. Stushnoff
Departments of Horticulture/Biochemistry, Colorado State University, Fort Collins, CO 80523, USA
D. L. Johnson
Department of Soil and Crop Sciences, Colorado State University, Fort Collins, CO 80523, USA
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 12 | Page No.: 2026-2029







ABSTRACT
Seasonal change in cold hardiness has frequently been associated with metabolic induction of osmolytes that may play a role in acclimation to low temperatures in cool season crop plants. The present research was aimed to examine the association of common endogenous sugars with cold hardiness in acclimated and non-acclimated plants of winter-hardy and non-hardy lentil (Lens culinaris Medik.) genotypes under controlled conditions. The genotypes were highly significantly (p<0.01) different for cold tolerance at -5°C and the winter hardy genotypes had no foliar damage but 75% injury was observed in the non-hardy genotypes when freeze tested after complete acclimation under controlled conditions. Comparatively, sorbitol concentrations in the tips and basal leaves of young and old shoots of acclimated plants were consistently and highly significantly (p<0.01) higher in the non-hardy cultivars than in the winter-hardy lines. A highly significant (p<0.001) positive correlation (0.74 ± 0.09) was found between cold injury and sorbitol concentration in these tissues. Except for the presence of comparatively high sorbitol in non-hardy genotypes, there was no significant association of other endogenous sugars nor with total soluble sugars and cold hardiness in lentil.
PDF Abstract XML References Citation
How to cite this article
Asghar Ali, C. Stushnoff and D. L. Johnson, 2000. Negative Association of Endogenous Sorbitol with Cold Hardiness in Lentil. Pakistan Journal of Biological Sciences, 3: 2026-2029.
DOI: 10.3923/pjbs.2000.2026.2029
URL: https://scialert.net/abstract/?doi=pjbs.2000.2026.2029
DOI: 10.3923/pjbs.2000.2026.2029
URL: https://scialert.net/abstract/?doi=pjbs.2000.2026.2029
INTRODUCTION
Lentil (Lens culinaris Medik), an important cool season food legume, is grown as a spring and winter-annual crop: Winter annuals can be acclimated under field or controlled conditions to tolerate rather low (-19°C) temperatures (Ali et al., 1991). Neither biochemical nor biophysical mechanisms controlling freezing tolerance in lentil have been studied previously. Understanding the mechanism controlling freezing tolerance in this crop may help breeders improve its productivity by manipulation of those factors responsible for cold hardiness. Total soluble carbohydrates and total sugars of aerial parts have been reported to be positively correlated with cold tolerance in different forage legumes (Acikgoz, 1982). In white clover (Trifolium repens L.), different genotypes from northern and southern parts of Norway were studied for carbohydrate concentration and it was found that soluble carbohydrates and sucrose contents increased in the stolons during hardening in the genotypes from both parts (Rosnes et al., 1993). Frost-hardy cabbage leaves had higher concentrations of sucrose and raffinose than non-hardy tissues and these concentrations decreased as dehardening proceeded (Santarius and Milde, 1977). Santarius (1973) showed that sugars protected thylakoids from freezing damage. Sugars were shown to accumulate in chloroplasts at low temperatures in cabbage, spinach (Krause et al., 1982) and wheat (Trunova and Zvereva, 1974). Fry et al. (1993) showed 47% more sucrose developed in growth chamber-acclimated centipede grass (Eremochloa ophiruoides H.) stolons compared to pre-acclimated stolons. A high positive correlation was reported between sucrose level and number of surviving stolons after exposure to freezing temperatures. In alfalfa (Medicago sativa), differences in the maximum degree of freezing tolerance between winter-hardy and non-hardy cultivars were more related to the capacity of the plants to accumulate stachyose and raffinose than to accumulate sucrose (Castonguay et al., 1995). In pea (Pisum sativum), snow tolerance was correlated with the cotyledon nonstructural carbohydrate concentrations which were 2-3 times higher than foliage non-structural carbohydrate concentrations (Fukuta and Yukawa, 1996).
On the other hand, no relationship, or an inverse relationship between carbohydrates and freezing tolerance has been reported by some researchers. Bula et al. (1956) found the highest concentrations of total soluble carbohydrates in the least hardy alfalfa cultivar. Sucrose concentration increased at the expense of starch during fall in bermudagrass (Cynodon spp.), but freezing tolerance was not affected (Dunn and Nelson, 1974). An inverse relationship between sucrose and raffinose concentrations and cold hardiness was reported in winter wheat (Green and Ratzlaff, 1975). Neither starch nor sucrose concentrations in stolons were related to freezing tolerance in Floratam St. Augustinegrass (Stenotaphrum secundatum K.) (Fry et al., 1991).
The objective of this study was to examine the association of common endogenous sugars to cold hardiness in lentil by analyzing acclimated and non-acclimated plants under controlled conditions.
MATERIALS AND METHODS
Sites and Plant Material: The experiment was conducted first at Colorado State University (USA) in 1994 and later repeated at National Agricultural Research Centre (Pakistan) in 1998. Initially, seven genetically stable lentil lines and varieties, including winter and spring lentils, were selected from our world collection (Table 1-4). Later, this number was reduced to four as a manageable experiment. Of these four lines, Balochistan Local (B. Local) and ILL 5865 (currently released as SHIRAZ-96) are known for their winter and cold hardiness while the other two, Brewer and Laird (spring cultivars), for their non-cold hardiness (Ali et al., 1991; 1999). All seven genotypes were used for analysis of random shoot (whole) samples of acclimated plants (Table 5). Initially, shoots were sampled randomly. Later, it was observed that young and old shoots had different cold tolerance limits. Therefore, shoot tips (leaf primordia) and basal leaves from old and young shoots were sampled to examine the differences in carbohydrate content and freezing tolerance among these tissues (Table 1).
Growth conditions, acclimation and freeze testing procedures: A well established protocol for screening of lentil for cold hardiness under controlled conditions was used for acclimation and freeze testing of the genotypes (Ali et al., 1999).
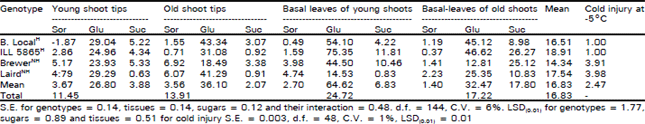
| Table 1: | Coacentrations (10–5 mol g–1) of sorbitol (Sor), glucose (Glu) and sucrose (Suc) in four different tissues of 6-week acclimated plants of two hardy (H) and non-hardy (NH) lentil genotypes and cold tolerance of the genotypes at -5°C freeze temperature where cold tolerance assessed on the basis of cold injury score (1 = no foliar injury to the plant and 5 = dead) |
 | |
| Table 2: | Correlation coefficients (± Standard Error) between cold injury, determined from a rating scale of 1-5 based on foliar injury and sugar content (10–5 mol g–1) in different acclimated plant samples of hardy and non-hardy lentil genotypes. Probability is in parenthesis |
| Table 3: | Concentrations (10–6 mol g–1) of sorbitol (Sor), glucose (Glu) and sucrose (Suc) in whole-shoot samples of non-acclimated-two hardy (H) and two non-hardy (NH) lentill genotypes |
 | |
| Table 4: | Concentrations (10–5 mol g–1) of sorbitol (Sor), glucose (Glu) and sucrose (Suc) in wholoolant (above-ground) samples of non-acclimated two hardy MI and two non-hardy (NH) lentil genotypes |
 | |
| Table 5: | Concentrations (10–5 mol g–1) of sorbitol (Sor), glucose (Glu) and sucrose (Suc) in whole-shoot samples of acclimated four hardy (H) and three non-hardy (NH) lentil genotypes |
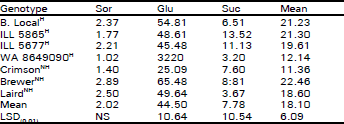 | |
S.E. for genotypes = 1.65, sugars = 1.08 and their interaction = 2.86. d.f. = 147. C.V. = 45%. NS = Non-significant (p>0.05) | |
Trays containing plants not to be exposed to acclitnation -treatments were placed in the greenhouse at 25±3°C day and 10±3°C night temperatures with a 12 h photoperiod (Table 3-4). The 6-week acclimated and control plants were first sampled and then tested for freezing tolerance by reducing growth chamber temperature at the rate of 1°C h–1 starting from 0°C to reach the minimum temperature of -5°C and then for 4 h holding at this temperature. The total time in freezer was 9 h for the plants tested. After freeze testing, plants were held at room temperature overnight and then transferred to the greenhouse at approximately 25°C with a 12 h photoperiod. Within 48 to 72 h of the freezing test, data for injury were recorded on the scale of 1 to 5 where 1 = no, foliardamage and 5 = dead (Ali et al., 1999; Table 1).
Sample preparation and ,derivetizetien: Plant samples were frozen in a freezer at --70°C or in liquid nitrogen (-196°C) and freeze-dried. All samples were ground and passed through a 400 mesh (US sievadesignation) stainless steel sieve prior to derivatization. A modified trirnethylsilation procedure (Sweeley et al., 1963) was used. -A-known quantity of 0.5 to 1.5 mg of ground sample was placed in a screw-capped vial, in duplicate, containing 25 μg of internal standard. Then 400 μl of pyridine, 80 μl of hexamethyldisilazane (98%) and 40 μ1 of trirnethylchlorosilane were added in- this order. The tightly capped tubes were heated at 80°C for 20 minutes. After heating, tubesAmere uncapped, blown dry at room temperature, and 0.5 ml-of-_n-hexane was added to the dried-residue in each. The solvent was pipetted into a clean, dry tube. The n-hexane solution was again blow dried and 200 μl of fresh n-hexane were added. One μl of this solution was injected with a dropping needle injector into gas chromatograph. Total four (Table 1, 3, 4) and eight (Table 5) samples were taken and each sample was run twice. The amount of each analyte was determined using the internal standard method. Sorbitol, glucose, sucrose, raffinose and stachyose were chromatographed on an PIP-1 methyl siloxane capillary column (12 m, 0.20 mm, 0.33 μm). Since raffinose and stachyose were completely absent or below the detectable concentration in all plant samples, they were not taken into account for any consideration or, presentation.
Statistical analysis: A completely randomized design (CRD, two or three-factor with four or eight replications) was used to analyze the data. Least Significant Difference (LSD) range test was used for mean separation. To determine association between carbohydrate concentrations and freezing ,tolerance, a simple correlation was run between concentration of sorbitol, glucose and sucrose and visible freezing injury in randomly sampled whole shoots and -the data from tips and basal leaves of young and old-shoots (Table 1, 2, 5).
RESULTS
The genotypes were highly significantly (p<0.01) different for cold tolerance at -5°C. The two winter hardy genotypes had no cold injury at this minimum temperature but 75% foliar damage was observed in the two winter non-hardy genotypes when freeze tested after complete acclimation under controlled conditions (Table 1). The control (non-acclimated) plants of all genotypes were completely killed at this temperature (data not presented). It shows the variability for freeze tolerance among the genotypes when acclimated.
Sorbitol concentrations in the tips and basal leaves of young and old shoots of fully acclimated plants were consistently and highly significantly (p<0.01) higher in the non-hardy cultivars than in the cold-hardy lines. There was no other trend found for other endogenous sugars in the genotypes (Table 1). A highly significant (p<0.001) positive correlation (0.74 ± 0.09) was found between cold injury and sorbitol in tips and basal leaves of young and old shoots of acclimated plants. There were no other significant (p>0.05) correlations between cold injury and other sugars in this study (Table 2). There were nonsignificant (p>0.05) differences for sorbitol among the genotypes when they were not hardened (Table 3, 4).
When whole shoots of acclimated plants were sampled at random, there were no significant (p>0.05) differences for sugar content among four hardy and three non-hardy genotypes. A high coefficient of variation was found for sugar content among genotypes (Table 5). In control (non-acclimated) plants, two non-hardy cultivars (Brewer and Laird) were significantly (p<0.01) higher in glucose and sucrose than one of the cold hardy lines (B. Local) but not higher than the other cold-hardy line (ILL 5865) (Table 3). Overall, there were no trends that suggested an association of endogenous sugars to cold hardiness when whole-shoots or whole-plants were sampled (Table 3-5).
DISCUSSION
After complete acclimation, the lentil plants show similar cold tolerance under controlled conditions as they show under natural environments (Ali et al., 1991; 1999). The cultivars or lines used in the study were pure, homozygous and genetically stable. The seeds were also obtained from the genotypes grown in the same environment. Furthermore, the plants were grown under controlled conditions where a minimum, micro-enviroment effect may be expected and a few tissues of the plant were sampled whereas the other similar tissues were left on the same plant for cold injury observations. Therefore, there are almost no chances of having any differences among the genotypes due to any environmental variation or handling the genetic material.
A positive correlation between cold injury and sorbitol was found indicating that higher sorbitol content was associated with lack of cold hardiness (Table 2). This is probably the reason that non-significant (p>0.05) differences for sorbitol were found among the genotypes when they were not hardened (Table 3, 4). But when hardened, these concentrations were highly significantly (p<0.01) higher in the non-hardy cultivars than the hardy lines and showed significant (p<0.05) differential response to freeze tolerance (Table 1). These data show that high concentrations of sorbitol are associated with lack of cold hardiness. It may be that during cold acclimation carbon resources are directed to some unknown protective metabolite in hardy tissues, but not in non-hardy tissues. It is also possible that sorbitol, in some unknown way, is associated with undesirable freezing events. The role of sorbitol in drought tolerance of faba bean (Vida faba L.), common bean (Phaseolus vulgaris L.) and chickpea (Cicer arietinum L.) has been reported by Amede and Schubert (1997). The concentration of sorbitol was substantially increased in faba bean during drought stress and the response of all species was accompanied by an increase in the solute pool which created a substantial decrease in osmotic potential and rate of cell division and cell expansion. The low temperature stress also affects the plant growth and in response to this environmental change, the winter hardy plants go under dormant conditions but winter non-hardy plants keep their growth continued and do not prepare themselves for the forthcoming undesirable events and hence die in the low temperatures (Levitt, 1972; 1980; Sakai and Larcher, 1987). One or both alternatives may be worthy of further consideration, but this study was not designed to explore these mechanisms. It does, however, suggest a starting point for further metabolic studies concerning a possible role of carbohydrate metabolism in cold survival of lentil.
A high coefficient of variation in whole-shoot samples suggested a high endogenous variability in sugar content among samples (Table 5). It is clearly evident that the metabolic changes in the plant body in response to environmental change are extremely important for its survival (Levitt, 1972; 1980; Sakai and Larcher, 1987). It is concluded that lack of cold hardiness was associated with increased concentration of sorbitol.
ACKNOWLEDGMENTS
The financial support of USAID and PARC under MART/AZR project for Ph.D. program and the research, work reported in this paper, and the seed provided by Dr. W. Erskine (ICARDA) and Dr. F. J. Muehlbauer (USDA, WSU) are gratefully acknowledged.
A part taken from thesis submitted to the Academic Faculty of Colorado State University in partial fulfillment of the requirements for the degree of Ph.D.
REFERENCES
- Ali, A., D.L. Johnson and C. Stushnoff, 1999. Screening lentil (Lens culinaris) for cold hardiness under controlled conditions. J. Agric. Sci., 133: 313-319.
Direct Link - Ali, A., J.D.H. Keatinge, B.R. Khan and S. Ahmad, 1991. Germplasm evaluation of dual-season lentil (Lens culinaris) lines for the arid highlands of West Asia. J. Agric. Sci., 117: 347-353.
CrossRefDirect Link - Amede, T. and S. Schubert, 1997. Solute Pool Enlargement in Drought-Sensitive or -Tolerant Grain Legumes: Concentration Effect or Osmotic Adjustment. In: Plant Nutrition for Sustainable Food Production and Environment, Ando, T., K. Fujita, T. Mae, H. Matsumoto, S. Mori and J. Sekiya (Eds.). Kluwer Academic Publishers, Dordrecht, Boston, London, pp: 165-66.
- Bula, R.J., D. Smith and H.J. Hodgson, 1956. Cold resistance in alfalfa at two diverse latitudes. Agron. J., 48: 153-156.
CrossRefDirect Link - Castonguay, Y., P. Nadeau, P. Lechasseur and L. Chouinard, 1995. Differential accumulation of carbohydrates in alfalfa cultivars on contrasting winterhardiness. Crop Sci., 35: 509-516.
Direct Link - Dunn, J.H. and C.J. Nelson, 1974. Chemical changes occurring in three bermudagrass turf cultivars in relation to cold hardiness. Agron. J., 66: 28-31.
CrossRefDirect Link - Fry, J.D., N.S. Lang and R.G.P. Clifton, 1991. Freezing resistance and carbohydrate composition of 'Floratam' St. Augustinegrass. HortScience, 26: 1537-1539.
Direct Link - Fry, J., N.S. Lang, R.G.P. Clifton and F.P. Maier, 1993. Freezing tolerance and carbohydrate content of low-temperature-acclimated and nonacclimated centipedegrass. Crop Sci., 33: 1051-1055.
CrossRefDirect Link - Green, D.G. and C.D. Ratzlaff, 1975. An apparent relationship of soluble sugars with hardiness in winter wheat varieties. Can. J. Bot., 53: 2198-2201.
CrossRefDirect Link - Rosnes, K., O. Junttila, A. Ernstsen and N. Sandli, 1993. Development of cold tolerance in white clover (Trifolium repens L.) in relation to carbohydrate and free amino acid content. Acta Agric. Scandinavica B-Plant Soil Sci., 43: 151-155.
CrossRefDirect Link - Santarius, K.A., 1973. The protective effect of sugars on chloroplast membranes during temperature and water stress and its relationship to frost, desiccation and heat resistance. Planta, 113: 105-114.
CrossRefDirect Link - Santarius, K.A. and H. Milde, 1977. Sugar compartmentation in frost-hardy and partially dehardened cabbage leaf cells. Planta, 136: 163-166.
CrossRefDirect Link