
Wan Mohtar Wan Yusoff
School of Biosciences and Biotechnology, Faculty of Science and Technology, University Kebangsaan Malaysia, 43600 UKM Bangi, Selangor, Malaysia
Muhannad I. Massadeh
School of Biosciences and Biotechnology, Faculty of Science and Technology, University Kebangsaan Malaysia, 43600 UKM Bangi, Selangor, Malaysia
Othman Omar
School of Biosciences and Biotechnology, Faculty of Science and Technology, University Kebangsaan Malaysia, 43600 UKM Bangi, Selangor, Malaysia
Jalil Kader
School of Biosciences and Biotechnology, Faculty of Science and Technology, University Kebangsaan Malaysia, 43600 UKM Bangi, Selangor, Malaysia
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 10 | Page No.: 1758-1761







ABSTRACT
Trichoderma reesei QM9414 was co-cultured with Aspergillus terreus SUK-1 for cellulase and reducing sugar production on sugar cane bagasse in solid substrate fermentation. The highest cellulase activity (FPase 0.68 (IU/ml), β-glucosidase activity (0.08 IU/ml) and reducing sugar concentration (1.4 mg/ml) were obtained in mixed cultures using inoculum ratio of 2:1 (Trichoderma : Aspergillus) compared to other ratios investigated (1:2 and 1:1). Nevertheless, experiments with pure culture of T. reesei was found to be better in terms of the production of FPase (1.4 IU/ml) and reducing sugar concentration (2.3 mg/ml), The addition of A. terreus to T. reesei in cultures of 2:1 inoculum ratio increases the production of β-glucosidase (the highest activity of 0.08 lUlml was detected in cultures of 2:1 ratio as well as in pure cultures of A. terreus). Nevertheless, this enhanced (β-glucosidase activity in mix culture could not result in enhanced substrate utilization and reducing sugar production. It was found that the highest percentage of degradation (25%) was achieved when pure cultures of T. reesei was employed followed by when pure cultures of A. terreus was employed (22%) and when mix cultures of 2:1 inoculum ratio was employed (22%). This suggests that synergism of β-glucosidase and cellulase to enhance substrate degradation was better affected using pure culture than mixed culture in solid substrate fermentation of sugar cane bagasse.
PDF Abstract XML References Citation
How to cite this article
Wan Mohtar Wan Yusoff, Muhannad I. Massadeh, Othman Omar and Jalil Kader, 2000. Sugar Cane Bagasse Degradation by Mixed Culture of T. reesei and A. terreus in Solid Substrate Fermentation. Pakistan Journal of Biological Sciences, 3: 1758-1761.
DOI: 10.3923/pjbs.2000.1758.1761
URL: https://scialert.net/abstract/?doi=pjbs.2000.1758.1761
DOI: 10.3923/pjbs.2000.1758.1761
URL: https://scialert.net/abstract/?doi=pjbs.2000.1758.1761
INTRODUCTION
Filamentous fungi grow typically in nature on solid substrates, such as wood, seeds, stems, roots and leaves of plants in symbiotic associations (Castillo et al., 1994). In the case of lignocellulose residues, the complexity of the substrate requires the action of multiple enzymes and the interaction of many microbes to accomplish biodegradation process. Mixed culturing of fungi on solid substrates thus may produce the optimal combination of enzymes for the most efficient biodegradation process (Gutierrez-Correa and Tengerdy, 1997). Although Trichoderma reesei is one of the best cellulase producers, its production of β-glucosidase is low (Ryu and Mandels, 1980) and therefore, may require the addition of β-glucosidase from other microbial source in a mixed culture fermentation. β-glucosidase enzyme is required to remove cellobiose which is a strong inhibitor for carboxymethylcellulase enzymes. Mixed culture of Trichoderma reesei and Aspergillus phoenicis in submerged fermentation was reported and in solid substrate fermentation (SSF), Duenas et al. (1995) reported that cellulase with an increased amount of β-glucosidase could be produced. Yusoff and Thayan (1991) reported that mix culturing of Trichoderma reesei and Aspergillus niger in submerged culture fermentation (SCF) resulted in good cellulase production with increased amounts of β-glucosidase enzyme but the synergism of β-glucosidase and cellulase from different species was lower than that observed in a pure culture of T. reesei. This paper is a following of the above report in mixed-fungal solid substrate fermentation (SSF) of sugar cane bagasse by T. reesei and A. terreus for cellulase production and substrate degradation.
MATERIALS AND METHODS
Microorganism and estimation of growth: The local isolate A. terreus 1M128243 and T. reesei QM9414 was obtained from the stock cultures of the Microbiology department (Universiti Kebangsaan Malaysia). The cultures were maintained on Potato Dextrose Agar (FDA) at 30°C for 3 days and then for a further 4 days at room temperature (27°C) before storing at 4°C. Spore suspension of 107 sporeiml was prepared by harvesting from 7-days old cultures of both molds with 10 ml sterile distilled water and 10% (vlv) of inoculum (of the tested ratios) was used in all experiments.
Substrate: Sugar cane bagasse. (without the hard outer layer) as the substrate and carbon source was obtained from night market area around Bandar Baru Bangi, Selangor. It was washed with distilled water (3 times), dried at 60°C and milled (National, MX-591 N) to 1-2 mm particle size. No chemical pretreatment was employed and only physical pretreatment by milling was carried out.
Medium: The medium employed (Sternberg, 1976) contains (NH4)2SO4, 1.4 g; KH2PO4, 2.0 g; CaCl2. 0.3 g; MgSO4, 0.3 g; FeSO4.7H2Q, 0.005 g; MnSO4.7H20, 0.0016 g; Zn504.7H2O, 0.0014 g; CoCl2, 0,002 g; protease peptone, 0.75 g and Tween 80, 1.0 g; per liter. The pH of the medium was adjusted to 5.5 at room temperature.
Solid substrate fermentation: Fermentations were conducted in 500 ml flasks containing 6 g dried milled sugar cane bagasse and 60 ml of Mandels medium. The flasks were autoclaved at 121°C for 15 min. Each flask was then inoculated with 10% (v/v) spore suspension (6.0 ml) of pure culture of Trichoderma reesei QM9414 or pure culture of Aspergillus terreus IM128243 (0:1) or mixture of both fungi (three different inoculum ratios were employed (2:1, 1:1 and 1:2; Trichoderma:Aspergillus). All flasks were incubated at 30°C for 8 days of fermentation. Duplicate flasks were set up for each experimental variation.
Sampling and Extraction method: Based on Muniswaran and Charyulu (1994), for each gram of initial substrate weight, 10 ml of distilled water was used to dispense the fermented moldy pith. The dispensed pith was shaken at 130 rpm at room temperature (27°C) far 1 hour. The mixture was filtered through nylon cloth of 200 mesh and the collected substrate was dried at 60°C for 24 hours to measure the percentage of degradation. The pH of the collected solution was measured before it was centrifuged. The supernatant was assayed for cellulase activity, reducing sugar and dissolved protein content.
The percentage of degradation was calculated as follows:
where, I. Wt is the initial weight of the substrate and F.Wt is the final weight after fermentation.
Analytical procedures: The endoglucanase activity was measured as carboxymethylcellulase (CMCase) using 3% carboxymethyl cellulose in 1.5 M Acetate buffer, pH 5.0, while for exoglucanase it was measured as filter paper activity (FPase) using Whatman No. 1 filter paper (Mandels et al., 1976). The absorbances were read using spectrophotometer (Hitachi 2000, Japan). β-glucosidase activity was assayed according to the method of Wood and Bhat (1988) using p-nitropheny1-6-D-glucopyranoside as substrate. Incubation of 1 ml of the enzyme with 1 ml of the substrate in 0.2M Phosphate buffer at pH 6.0 for 20 minutes at 37°C. The activities are expressed as international units (IU), defined as the amount of enzyme required to produce one pmole of glucose or p-nitrophenol (in case of β-glucosidase) per minute. The amount of reducing sugars released by the hydrolysis of bagasse was measured using Nelson-Somogyi method (Nelson, 1944; Smogyi, 1952) and the dissolved protein amount was measured employing Lowry method using bovine serum albumin as standard (Lowary et al., 1951).
RESULTS AND DISCUSSION
The results shown in Fig. 1-3 indicated the evaluation of the effects of different inoculum ratios on cellulose hydrolysis in SSF cultures based on production of reducing sugar, FPase and β-glucosidase activities.
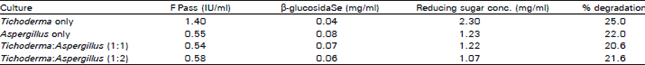
The results in Fig. 1 indicate that pure culture of Trichoderma gave the highest activities of FPase compared to other ratios employed. The highest values for FPase activity detected was 1.4 IU/ml (after 6 days of fermentation) followed by cultures using 2:1 ratio (0.68 IU/ml), cultures using 1:2 ratio (0.58 IU/ml), pure cultures of Aspergillus (0.55 IU/ml) and cultures using 1:1 ratio (0.54 IU/ml) after the same time of fermentation. The competition for growth or a lack of synergism of the enzymes between the low cellulase producer (Aspergillus) and the high cellulase producer (Trichoderma) may explain the reduced cellulase activity of the mixed culture (Table 1).
As A. terreus did not produce as much reducing sugar compared to T. reesei, it must rely on cellulase hydrolysis by Trichoderma to obtain reducing sugar for its growth (Duenas et al., 1995). Reducing sugar production reduces by 50% in mix cultures compared to when pure culture of Trichoderma was employed. The sugar consumption by the Aspergillus relieves the inhibition effect by sugar and resulted in a higher β-glucosidase production. Reducing sugar profile also showed similar production profile (i.e., reduction in reducing sugar concentration in mixed cultures) compared to when pure culture of Trichoderma was employed. Cultures using Trichoderma only gave the maximum reducing sugar concentration (Fig. 2) compared to other cultures. The results obtained when pure culture of Aspergillus were employed showed that Aspergillus has the ability to degrade sugar cane bagasse without any enzyme contribution from Trichoderma. This is supported from the production of reducing sugar detected, albeit less than 1 mg/ml. Whereas, in pure culture of Trichoderma, the results reflect its ability to degrade sugar cane bagasse best and are supported by the highest reducing sugar concentration detected. This leads to claim that the best mixed culture for hydrolysis could not match the hydrolytic potential of enzymes from pure cultures especially in this case that of Trichoderma.
The data shown in Fig. 3 indicates that the addition of A. terreus to Trichoderma reesei enriches the cultures with β- glucosidase enzyme. The maximum β-glucosidase activity of 0.08 IU/ml was obtained in pure culture of Aspergillus followed by mixed culture 2:1 compared to other cultures employed. Nevertheless, the increased β-glucosidase activity in mix cultures did not result in increasing the reducing sugar production compared to pure cultures of Trichoderma reesei with a 50% lower β-glucosidase activity. All these results are in agreement with results reported by Yusoff and Thayan (1991) where they claim that β-glucosidase from Trichoderma though having the lowest activity compared to that in mixed cultures (Submerged culture fermentation) has higher hydrolytic potential or synergism. Our results show that Trichoderma reesei produces a better hydrolytic enzyme mix and have the highest synergy for the hydrolysis of sugar cane bagasse in SSF compared to pure cultures of Aspergillus terreus or mixed cultures of both fungi. Table 1 contains data that further supports the previous claim, that is a higher substrate degradation (of 25%) was achieved when pure cultures of Trichoderma reesei was employed compared to 22% degradation by mix cultures of the ratio 2:1 (Trichoderma : Aspergillus) also with enzymes contribution from Trichoderma. The growth was estimated indirectly by measuring the dissolved protein amount in each sample (Fig. 4). The growth rate of cultures containing Trichoderma alone was found to be the highest. Whilst, in mixed cultures where Aspergillus was in competition with Trichoderma, the growth was poor and lower values of proteins were detected compared to experiment using pure cultures of Trichoderma. The origin of β-glucosidase seems to be very important in establishing an operating ratios of.
Fpase: β-glucosidase required for the effective hydrolysis of cellulosic substrates (Ryu and Mandels, 1980). The cellulase itself is a complex enzyme that contains many components that varied from one microorganism to another. Therefore, more work needs to be undertaken, may be at a molecular level to further elucidate the mechanism of synergism. The pH profile shown in Fig. 5 indicated cellulase and reducing sugar production in pure and mix cultures are generally within the same pH time profile where it can be seen that the final pH for pure cultures of Trichoderma reesei was recorded in the range of 3.1-3.3 after the first 48 hours. The other cultures (1:1, 1:2 and 2:1) shows similar pH profiles after 48 hours of fermentation and the pH was in the range of 3.2-3.8 with the exception of Aspergillus terreus where the pH range was 3.2-4.1 in the same period of fermentation.
| Table 1: | Results of maximum celluiase activities (IU/ml) and reducing sugar concentration obtained in pure and mixed culture SSF of Trichoderma ressei QM9414 and Aspergillus terreus IMI28243 |
 | |
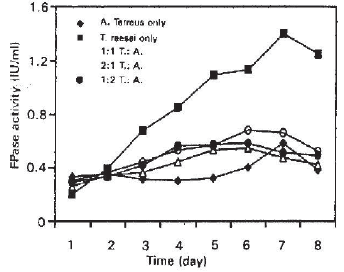 | |
| Fig. 1: | Fpase activity profiles from pure and mixed solid substrate fermentation of Trichoderma reesei QM9414 (T.) and Asepergillus terreus IMI28243 (A.) |
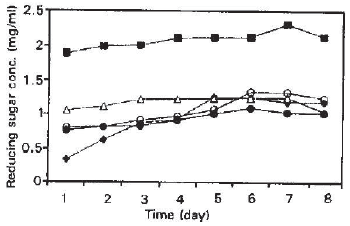 | |
| Fig. 2: | Reducing sugar concentration from pure and mixed solid substrate fermentation of Trichoderma reesei QM9414 (T.) and Asepergillus terreus IMI28243 (A.) |
in conclusion, our results suggested that the hydrolytic potential of cellulolytic enzymes could not be enhanced synergistically by supplementing components of enzyme (like β-glucosidase) from a different microorganism but rather should be from the same microorganism. The conclusion holds true in SCE and SSF mixed culture fermentation of sugar cane bagasse.
ACKNOWLEDGMENT
The authors greatly acknowledge a grant from the Ministry of Science, Malaysia, IRPA 08-02-02-001 for their financial support.
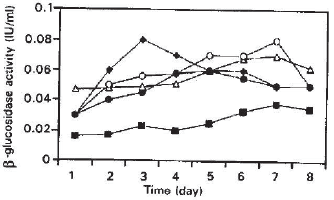 | |
| Fig. 3: | β-glucosidase activity profiles from pure and mixed solid substrate fermentation of Trichoderma reesei OM9414 (T.) and Asepergillus terreus IMI28243 (A) |
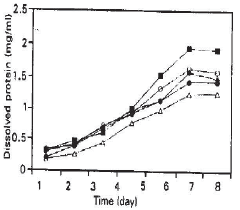 | |
| Fig. 4: | Dissolved protein concentration from pure and mixed solid substrate fermentation of Trichoderma reesei QM9414 and Aspergillus terreus IMI28243. Symbols are the same as in Fig. 1 |
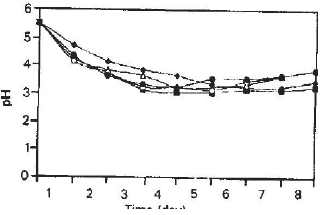 | |
| Fig. 5: | pH profiles of pure and mixed solid substrate fermentation of Trichoderma reesei QM9414 and Aspergillus terreus IM128243. Symbols are the same as in Fig. 1 |
REFERENCES
- Castillo, M.R., M. Gutierrez-Correa, J.C. Linden and R.P. Tengerdy, 1994. Mixed culture solid substrate fermentation for cellulolytic enzyme production. Biotechnol. Lett., 16: 967-972.
CrossRefDirect Link - Duenas, R., R.P. Tengerdy and M. Gutierrez-Correa, 1995. Cellulase production by mixed fungi in solid-substrate fermentation of bagasse. World J. Microbiol. Biotechnol., 11: 333-337.
CrossRefDirect Link - Gutierrez-Correa, M. and R.P. Tengerdy, 1997. Production of cellulase on sugar cane bagasse by fungal mixed culture solid substrate fermentation. Biotechnol. Lett., 19: 665-667.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Mandels, M., R. Andreotti and E. Roche, 1976. Measurement of saccharifying cellulase. Biotechnol. Bioeng. Symp., 6: 21-33.
PubMed - Muniswaran, P.K.A. and N.C.L.N. Charyulu, 1994. Solid substrate fermentation of coconut coir pith for cellulase production. Enzyme Microb. Technol., 16: 436-446.
CrossRefDirect Link - Nelson, N., 1944. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem., 153: 375-380.
Direct Link - Ryu, D.D.Y. and M. Mandels, 1980. Cellulases: Biosynthesis and applications. Enzyme. Microb. Technol., 2: 91-102.
CrossRefDirect Link - Sternberg, D., 1976. Production of cellulase by Trichoderma. Biotechnol. Bioeng. Symp., 6: 35-53.
PubMed