
Raees Hussain Zaidi
Department of Zoology University of Karachi, Karachi-75270, Pakistan
S. Shahid Shaukat
Department of Botany, University of Karachi, Karachi-75270, Pakistan
Shaheen Abidi
Department of Zoology University of Karachi, Karachi-75270, Pakistan
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 1 | Page No.: 126-129







ABSTRACT
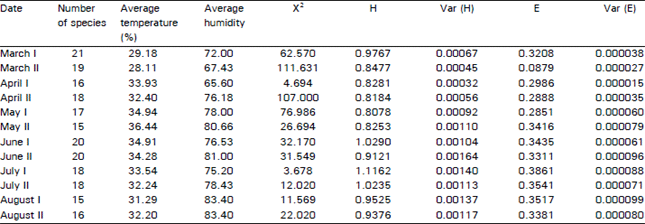
A tropical insect community was analyzed from March to August, 1990. The observed abundance pattern usually did not fit MacArthur's broken stick model. Species diversity (H’) was highest in June and July (1.0236, 1.0290 and 1.01162) and lowest in April and May (0.8078 and 0.8281). Agglomerative cluster analysis showed two groups of samples with the exception of samples collected in May. First group consisted of March to April samples and 2nd group had the samples of May to August. This clustering revealed that community changes were correlated with the environmental factors i.e. humidity, temperature and rainfall.
PDF Abstract XML References Citation
How to cite this article
Raees Hussain Zaidi, S. Shahid Shaukat and Shaheen Abidi, 2000. Multivariate Analysis of a Tropical Insect Community. Pakistan Journal of Biological Sciences, 3: 126-129.
DOI: 10.3923/pjbs.2000.126.129
URL: https://scialert.net/abstract/?doi=pjbs.2000.126.129
DOI: 10.3923/pjbs.2000.126.129
URL: https://scialert.net/abstract/?doi=pjbs.2000.126.129
INTRODUCTION
The insect community studied was an admixture of various insect groups i.e., ants, bugs, beetles, butterflies, demsel flies, dragon flies, moths treehoppers and spiders. Most of members of this community infest Albizia lebbeck, Acacia nilotica and prosopis julisflora (Ahmad and Abrar, 1976; Zaidi et al., 1991). The present paper analyses the community dynamic of the insect community at Karachi University Campus. Callan and Cranston (1994) and Howse et al. (1998) studied population of various insect groups. Zaidi et al. (1991) studied an Oxyrhachis dominated community and examined community dynamics through examined the structure of communities of different multivariate techniques of data analysis. The present paper the structure, composition and the environmental relationships of a tropical insect community.
MATERIALS AND METHODS
Sampling: An open desert plant community situated at Karachi University Campus was selected for the present studies. insect fauna was collected from March to August 1990 at 15 days intervals by beating and sweeping methods using a net with a radius of 14 inches. The net was placed under the branches and beaten three times in each sample while collection from smaller plants and grasses were made by sweeping for which purpose a quadrat of size 2×2 mm was randomly selected. These methods were also used by Richards and Waloff (1961), Legner and Oatman (1962), Gibbs and Betts (1963), White (1675) and Ahmad and Zaidi (1985) for various insects Ants. bugs, beetles, butterflies, damsel flies, dragon flies, dusky cotton bugs, moths, treehoppers and spiders were collected and stored separately. Relative humidity and temperature within the community were recorded during the sampling period.
Species diversity: Species richness is simply expressed as the number of species S. The general diversity is computed using Margalef (1958) information theory index (H’) as follows:
Where ni is the number of individuals of with species and N equals the total number of individuals in the sample. The variance of H' is computed as:
Equitability is expressed by Pielou (1975) index:
E’ = H/H max = H’/log (S)
The variance of E’ is as follows:
Var (E’) = Var (H’) / (log S)2
The relative abundance pattern of species in the community at various time intervals is portrayed in the form of species rank/relative abundance plots (Whittaker, 1965; May, 1975). The structure of the community was analyzed in greater detail by fitting MacArther’s broken stick model design posed by this model. The expected abundance of a species in broken stick model is:
Multivariate analysis: Principal component analysis (PCA) was performed to seek trends in the data structure while cluster analysis was accomplilshed using minimum variance clustering (Orloci, 1978) to detect group structure.
RESULTS AND DISCUSSION
Species diversity: In general species diversity was low. Both species richness and equitability components of diversity were generally low and varied over the observation period (Table 1). Highest diversity was recorded in June and July and lowest in April and May. Diversity was uncorrelated with temperature but showed some relationship with humidity and rainfall. Diversity tends to increase with increasing humidity and with the rainfall. The rank order/abundance curves are given in Fig. 1, In most of the samples diversity pattern is similar with each graph being in the form of a slightly curved line. Only one sample (second week of July) showed a close fit to MacArthur's broken stick model while the rest of the samples departed significantly from this model (Table 2) (p<0.001).
| Table 1: | Species richness (S), diversity (H), equitability (E) and X2 value for goodness of fit with MacArthur's broken stick model. I. First fortnight; II, second fortnight |
 | |
| Table 2: | Results of principal component analysis showing eigen values, explained various eigen vector elements and the associated variables (species) |
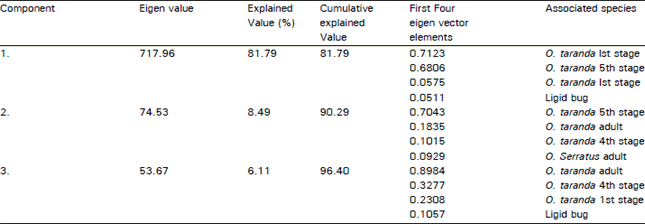 | |
Ordination: Fig. 2 shows ordination of samples A seasonal trend is readily seen in the three dimensional configuration. The samples collected during March to May fall in the lower right of the ordination while those of June to August are situated in the upper left of the configuration. The first ordination axis which explained 81.75 percent of variation showed significant correlation with humidity (r = 0.714, p<0.01) and with temperature (r = 0.638, p<0.05). The second axis that explained 8.45 percent of total variability was significantly correlated with humidity (r =0.845, p<0.001) while the third axis that explained 6.11 percent of total variation was uncorrelated with the environmental factors.
Cluster analysis: Fig. 3 shows the result of agglomerative cluster analysis in the form of a dendrogram. Two broad groups can easily be recognized. Group I includes samples from March to May (1st fortnight) and the other groups (group II) includes samples from May (2nd fortnight) to August. Diversity was lower for group I compared to group II.
Average temperature for group I was 31.822°C while that for groups II was 33.55°C while average humidity was 71.84 and 79.94 percent respectively for group I and II. I. Taranda and O. Serratus were relatively less abundant in group I compared to group II while dusky cotton bug and ed cotton bug were relatively more abundant in group I and less abundant in group II. Rest of the species (spider, beetles, ligids, dragon fly and demsel fly) did not show any substantial variation in relative abundance with respect to season. Ahmad and Abrar (1976) reported that O. Taranda and O. Serratus are found on the same host are abundant in April and May. During present studies population of both species increased gradually from March to April and reduced with the same pattern from May to August. Earlier studies of Ahmad and Abrar (op. Cit.) Reported that these two species are sympatric. It was also observed that species abundance of most species increased from March to first half of May and decreased from 2nd half of May to August.
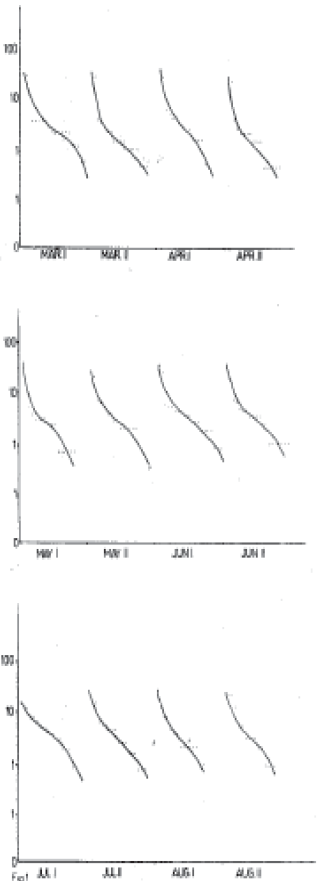 | |
| Fig. 1: | The rank order/abundance curves for the insect assemblages dated March 1 to August 2. Sequence of communities as in Table 1. |
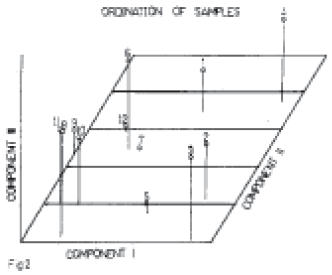 | |
| Fig. 2: | The principal component ordination of the insect assemblages |
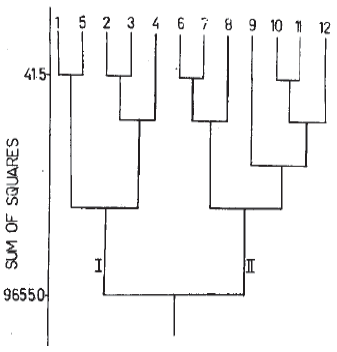 | |
| Fig. 3: | Dendrogram of the insect assemblages derived from agglomerative cluster analysis |
Average temperature of March-May is 31.82°C and May-August is 33.52°C. Ahmad and Zaidi (1985) and Jackson and Patana (1980) reported that temperature plays as important role in the biology of insects. Jackson and Patana (op. cit.) Reported that, development time of pink bollworm, Pectinophora gossypiella (Saunder) from 18.0±0.7 days at 20°C and it was 7 reduced to 6.8±0.5 days at 35°C. Shaukat and Khan (1990) also reported that grasshopper population was abundant in high temperature. Culin and Yeargan (1983) reported that in spider community abundance was maximum in August on soybean but the population of afa-alfa was maximum in December and thereafter decreased gradually. This variation was described by Turnbull (1973) who suggested that alfa-alfa provides a suitable over wintering site for spiders.
REFERENCES
- Ahmad, I. and R.H. Zaidi, 1985. Horizontal and vertical estimates of immatures survivorship in relation to population dynamics of red cotton stainer Dysdercus koeniggi (Fabr.) Herniptera: Pyrrhocoridael at Karachi University Campus. Proceedings of the 6th Pakistan Congress of Zoology, December 22-24, 1985, Zoological Society of Pakistan, pp: 141-148.
- Culin, J.D. and K.V. Yeargan, 1983. Comparative study of spider communities in alfalfa and soybean ecosystems: Foliage-Dwelling spiders. Ann. Entomol. Soc. Am., 76: 825-831.
CrossRefDirect Link - Legner, F.E. and E.R. Oatman, 1962. Foliage-feeding Lepidoptera on young nonbearing apple trees in Wisconsin. J. Econ. Entomol., 55: 552-554.
CrossRefDirect Link - Turnbull, A.L., 1973. Ecology of the true spiders (Araneomorphae). Annu. Rev. Entomol., 18: 305-348.
CrossRefDirect Link - White, T.C.R., 1975. A quantitative method of beating for sampling larvae of Selidosema suavis (Lepidoptera: Geometridae) in plantations in New Zealand. Can. Entomol., 107: 403-412.
CrossRefDirect Link