
Pear Mohammad
The United Graduate School of Agricultural Sciences, *Faculty of Agriculture, Ehime University, 3-5-7 Tarumi, Matsuyama 790-8566, Japan
Masaya Shiraishi
The United Graduate School of Agricultural Sciences, *Faculty of Agriculture, Ehime University, 3-5-7 Tarumi, Matsuyama 790-8566, Japan
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 2 | Page No.: 575-582







ABSTRACT
The anatomy of young roots of trifoliate orange budded with satsuma mandarin cultured hydroponically under pH 4.5-5.6, 6.0-6.3 and 7.5-7.6 were investigated to know the pH optima for this plant. Under pH 4.5-5.6, root tips were pointed and meristematic zone appeared wider. Starch grains were found on the rootcap and cortical cells apart from the tip but were absent completely from the cortical cell files close to the root apex. These characteristics remained similar along the exposure time. Under pH 6.0-6.3, roots were subjected to gradual shortening of growing zone followed by epidermal and root tip injuries on longer induction. Starch grain content in the cortical cells gradually increased up to 10 days of induction followed by conspicuous decrease, especially near the injured epidermis. The adverse effects of pH 7.5-7.6 started at short exposure resulting the severe sloughing of rootcap cells and bulbous structure of root tip. The epidermal injury also became conspicuous even within 3 days. The root apex secreted mucilaginous substances which were followed by injuries. Numerous starch grains were initially accumulated in the cortical cells but degraded completely at longer exposures. The cortical cells remained elongated and cell length gradually increased from the root apex towards the root base at pH 4.5-5.6. In contrast, the cells turned to round and their lengths became higher near the root apex in pH 6.0-6.3 and 7.5-7.6 compared to pH 4.5-5.6.
PDF Abstract XML References Citation
How to cite this article
Pear Mohammad and Masaya Shiraishi, 1999. pH Optima for Citrus Inferred from the Anatomical Records of Young Roots of Trifoliate Orange Budded with Satsuma Mandarin. Pakistan Journal of Biological Sciences, 2: 575-582.
DOI: 10.3923/pjbs.1999.575.582
URL: https://scialert.net/abstract/?doi=pjbs.1999.575.582
DOI: 10.3923/pjbs.1999.575.582
URL: https://scialert.net/abstract/?doi=pjbs.1999.575.582
INTRODUCTION
The effects of growing media pH on solubility, uptake and function of various ions as well as plant growth are well documented and widely anticipated (Feigin et al., 1986). On the other hand, the direct hydrogen-ion toxicity was described as difficult to demonstrate because the increase of ion was accompanied by many changes in the substrate which indicated that there might have some indirect effects of media acidity on plant function (Smith, 1971). However, although numerous reports on the effects of growing media pH have been published in different crops (Medappa and Dana, 1970; Zieslin and Snir, 1989; Rosen et al., 1990; Brunet, 1994; Sasada et al., 1994; Zieslin and Abolitz, 1994; Koyama et al., 1995), majority of these were broadly related to the nutritional or physiological phenomenon in plants. The above ground parts of plants deserved unique priority for these studies and under ground parts were usually unheeded. Exceptionally, although both above and under ground parts were subjected to these kinds of studies in citrus, the effects of media pH were even then remained unsolved with different inconsistent findings. Due to these anomalies, the pH optima for citrus is in contention.
Chapman (1968) reviewed that a wide range of media pH between 3.8-9.7 were tolerated by citrus plants. Other complementary studies revealed that although citrus plants endured a wide range of pH, all the ranges were not equally favored (Smith, 1971; Yokomizo and Ishihara, 1973). Therefore, it might be a logical hypothesis that plants exposed to different levels of growing media pH will response variously to the chemical factors involved. In order to have an increased understanding on these effects and to infer the optimum range of pH for citrus, an investigation on the matter at the tissue and cellular levels is required. In this connection, it is known that root growth is a result of two processes, cell division and cell expansion (Mayber and Lerner, 1994). These two processes are independent but sequential and must be coordinated for growth to occur. However, the length of developing cells have been suggested as the indicator of growing state of root (Silk et al., 1989). It has also been recently shown that growing cells do not necessarily respond to environmental stresses in the same way as the mature tissue (Nonami and Boyer, 1989; Pritchard et al., 1991). Thus determination of growth parameters in mature tissue does not accurately reflects controls of growth, since the site of measurement and the site of growth are spatially separate. For this reason, only the investigations on growing tissue or cells are important to ascertain the favorable environmental condition.
This study was intended to investigate the pH optima for citrus observing the effects of different culture solution pH on the anatomy of roots of trifoliate orange budded with satsuma mandarin with special emphasis on the root tip condition as well as the morphology of epidermal statures and the state of starch grains in the cortical cells, The cell length in the cortical files close to the root apex were also taken into account.
MATERIALS AND METHODS
The experiment was conducted in the Citriculture Laboratory, Faculty of Agriculture, Ehime University, Japan from April 14 to June 30 of three consecutive years (1996-1998) using three-year-old satsuma mandarin (Citrus unshiu Marc. cv. Okitsu Wase) trees grafted on trifoliate orange rootstocks. Eighteen trees for uniformity of size were selected and placed into styrofoam boxes retaining nutrient solution. Two trees were allocated for one box and the boxes were buried into the soil. The solution pH of all boxes was adjusted primarily to 5.0 everyday. The nutrient solution contained N, Ca, K, S, Mg, P, Na, Cl, Mn, Fe, B, Mo, Zn and Cu at approximately the following ppm, respectively: 198, 160, 80, 38, 23, 15, 11, 1, 0.5, 0.4, 0.25, 0.1, 0.05 and 0.02 as was used by Smith (1971). On the formation of numerous new roots, they were marked with wax-coated paper tags. Three boxes retaining six trees were subsequently considered as one treatment. Three levels of pH (4.5, 6.0 and 7.5) as three treatments were adjusted by titration of the nutrient solution with 1 M H2SO4 or 3 M NaOH as were required. The solutions had a tendency for the pH to drift up to 5.6, 6.3 and 7.6, respectively in every 24 hours thus the treatment turned to pH 4.5-5.6, 6.0-6.3 and 7.5-7.6. The pH of the nutrient solutions in every treatment were therefore adjusted daily to keep the pH near the desired levels. The nutrient solutions were completely renewed at 2 weekly intervals to ensure an adequate supply of all essential elements to the trees. The nutrient solutions were also continuously aerated and stirred by air pumps throughout the experimental period.
After 5, 10, 15 and 20 days of placement of trees to the treatments, roots were collected and washed gently several times with water. Segments of 6 mm length from the root apices were fixed immediately in 4 percent gluteraldehyde, buffered to pH 7.4 in sodium cacodylate, dehydrated in a graded ethanol series, infiltrated and embedded in JB-4 resin. Median longisections of three-micrometer thickness were stained with iodine-potassium-iodide. The sections were viewed and photographed under light microscope. Five roots of each treatment and nearly 30 sections from each root were examined every year. The photographs were subsequently studied for their comparison. Data on cell lengths were obtained from prints with a total magnification of x 1000 as was used by Bertaud and Gandar (1985). The same files were used for the measurements. Cell length at each point was taken as an average of 25 to 35 cells. The three-year-observations were interpreted and presented.
RESULTS
Anatomical features with special emphasis on root tip: The 5-day-induced roots showed pointed tips and received no root tip injuries or abnormalities under pH 4.5-5.6 (Fig. 1A). The meristematic region was clear which represented also a wide and well-developed column of cells to furnish the rootcap zone front ward and the sound cell files towards the root base. The quiescent center was far apart from the root tip. Ten-day-induced roots also had pointed tips without any injuries (Fig. 1B). All characteristic features resemble the roots under 5 days of induction. Meristematic region remained broader as was in the 5-day-induced roots, Similarly the 15 and 20-day-induced roots showed normal anatomy of all cell files (Fig. 1C, D). The cortical cells near the root apex were elongated but smaller and remained elongated with the increase in length towards the root base. In contrast, the 5-day-induced roots under pH 6.0-6.3 displayed apparently normal anatomy (Fig. 2A). Rootcap was centrally furnished with well-developed column of cells and meristematic region was wider. The quiescent center was far apart from the root tip resembling the normal root. The 10-day-exposed roots had the sloughing of rootcap cells and the quiescent center became proximal to the root tip (Fig. 2B). The growing zone shortened compared to pH 4.5-5.6. Epidermal injuries initiated in few places and root tip injuries were observed in 15-day-exposed roots. As a result, the meristematic region was partially destroyed (Fig. 2C). The cortical cells near the root tip became round. After 20 days, the root tip injury became more prominent destroying completely the quiescent center (Fig. 2D). The growing zone was totally changed by the presence of rounded cells. The epidermal cells were also under injury induction process.
Discharging of mucilaginous substances in the tip region became distinct in 5-day-induced roots under pH 7.5-7.6 (Fig. 3A). The rootcap cells sloughed off and gave the root tip dome shape. The meristematic region was markedly shortened producing rounded cells and root tip injuries also initiated. Epidermal injuries were started in few places. Tenday-induced roots under this treatment were subjected to distinct root tip injury and severe removal of rootcap cells after the secretion of mucilaginous substances (Fig. 3B). The root tip injury resulted the partial losing of the meristematic region. The cell rounding happened even within the surrounding cells of quiescent center. After 15 days, root tips became bulbous with the complete losing of rootcap and more removal of the growing zones (Fig. 3C). The cell rounding and enlargement continued and apical growing zone completely destroyed. Epidermal injuries and its destruction became more clear. After 20 days, the root tip injuries proceeded more towards the root base destroying the apical meristem greatly (Fig. 3D). The cell rounding was in progress.
Starch grain accumulation and degradation pattern: Under pH 4.5-5.6 starch grains accumulated on the rootcap cells, especially on the outer layers of rootcap region (Fig. 1A-D). The cortical cells were devoid of grains near the root apex (Fig. 1A-D). While the cortical cells of about 3-5 mm back from the root apex possessed a few starch grains (SG) near the epidermis (Fig. 4A-D). The trend of this accumulation presumed to be similar in all exposures. Under pH 6.0-6.3 starch grain accumulation and degradation pattern varied with exposure time. However, after 5 days induction, the rootcap cells contained a few starch grains on the outer
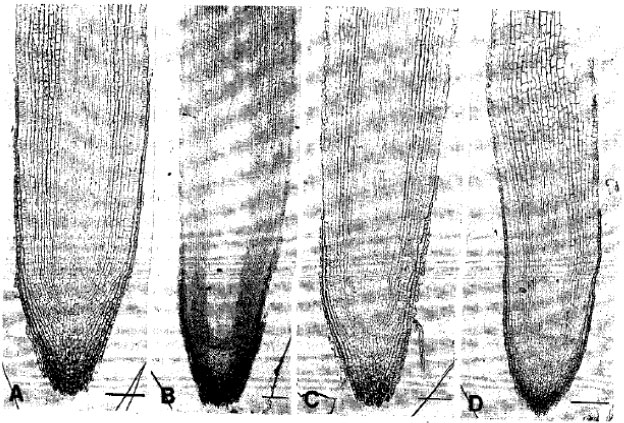 | |
| Fig. 1: | Light micrographs of roots of trifoliate orange budded with satsuma mandarin under pH. 4.5-5.6. A: Five-day induced B: Ten-day-induced C: Fifteen-day-induced D: Twenty-day-exposed. Note: Overall normal anatomy along the exposure time. Bar = 150 μm |
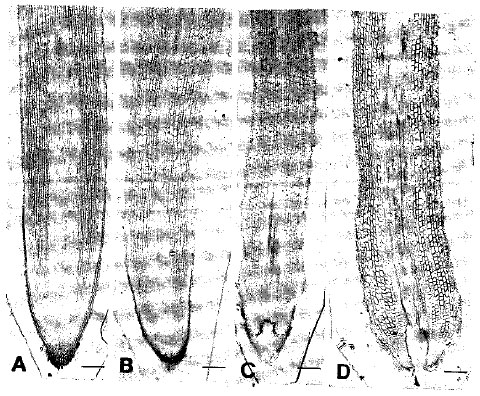 | |
| Fig. 2: | Light micrographs of roots of trifoliate orange budded with satsuma mandarin under pH 6.0-6.3. A: Five-day-induced with normal anatomy. B: Ten-day-induced. Note: Severe removal of rootcap cells and shortening of growing zone. C: Fifteen-day-induced. Note: Root tip injuries. D: Twenty-day-exposed. Note: Severe root tip injury. Bar 150 μm |
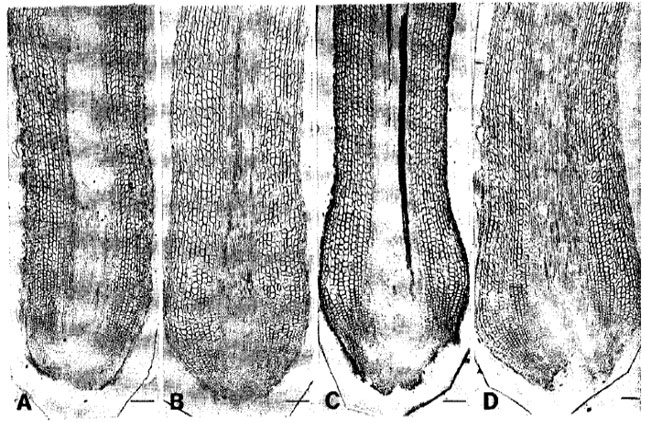 | |
| Fig. 3: | Light micrographs of roots of trifoliate orange budded with satsuma mandarin under pH 7.5-7.6. A: Five-day-induced with sloughing of rootcap and shortening of growing zone. B: Ten-day-induced having severe removal of root cells and shortening of growing zone. C: Fifteen-day-induced receiving severe root tip injury. D: Twenty-day-induced having destruction of root tip and shortening of growing zone. Bar = 150 μm |
layers (Fig. 2A). The cortical cells apart from the root apex accumulated comparatively much grains (SG) (Fig. 4E). The 10-day-induced roots also contained a few grains on the rootcap cells (Fig. 2B). Many starch grains (SG) were deposited on the cortical cells (Fig. 4F) but the degradation of grains near the injured epidermis (El) were noticed (Fig. 4F). On the 15th day starch grains (SG) were found on the cortical cells but were smaller compared to 10-day-induced roots (Fig. 4G). The grains near the injured epidermis (El) remained under degradation (Fig. 4G). On the 20th day the cortical deposition of starch grains (SG) were lesser and degradation of grains near the injured epidermis (El) became more conspicuous (Fig. 4H). Under pH 7.5-7.6, starch grain (SG) deposition became massive on the 5th day of induction on the cortical cells along with the degradation of grains near the injured epidermis (Fig. 4I). The deposition of grains (SG) became lesser on the 10th day and their degradation near the injured epidermis became clear (Fig. 4J). Starch grains were very less on the 15th day and degraded completely on the 20th day of induction (Fig. 4K, L).
Cell length in the cortical files close to the root apex: The cell length profile over the growing zone of roots under various treatments differed considerably in corresponding locations of files. However, the cell length in the roots under pH 4.5-5.6 was very less near the apex and gradually became larger towards the root (Fig. 5). This trend remained similar even after long exposures. Up to 1 mm from the root apex, cell length indicated that the cells were in dividing stage as the lengths remained below 20 μm. In contrast, the cell length in pH 6.0-6.3 treatment became comparatively bigger than the root apex although were smaller than the distal call length (Fig. 6). The cell length exceeded 20 μm even surrounding the quiescent center. This cellular elongation proceed more closer to the root apex along the exposure time. The induction period increased, the cell length also increases in the similar location, especially near the quiescent center under this treatment. Under pH 7.5-7.6, the cells near root apex modified to more longer even within slow exposure (Fig. 7). The quiescent center adjacent cells became larger. This trend gradually increased along with induction period. However, in every cases, the distal cortical cells were similarly enlarged.
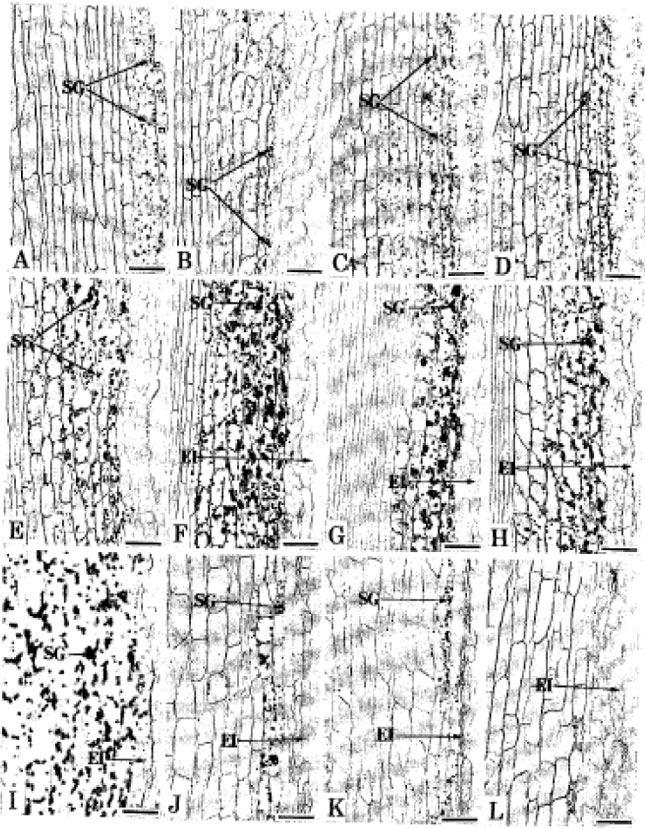 | |
| Fig. 4: | |
DISCUSSION
The normal growing state of roots under pH 4.5-5.6 probably utilized the maximum synthesized products for their nourishment and did not accumulate much starch grains in the usually stored sites of roots i.e. in the cortical cells. Starch grain accumulation on the cortical cells was reported under cold stress condition which. were utilized with the resumption of favorable temperature (Mohammad and Shiraishi, 1998). Therefore, our assumption on the normal stage of roots and the utilization of stored products by them under pH 4.5-5.6 were supported by this report. The intact roots with elongated tips also stood in favor of the conclusion that pH 4.5-5.6 provided better condition to the plants. Recently in another report pH 4.5-5.6 was also suggested as the better range for this plant in terms of root formation, root growth and root morphology (Mohammad and Shiraishi, 1999). Our present anatomical study also suggested this range as better than pH 6.0-6.3 and 7.5-7.6.
Injuries in 10-day-induced roots under pH 6.0-6.3 perhaps indicated that these roots were under adverse condition in this treatment. The complete abolition of apical regions through root tip and epidermal injuries along with starch grain degradation around the injured portions also suggested this treatment as adverse. The observance of much starch grains on the cortical cells in the preliminary stage might be indicative that growth cessation started shortly and degradation of grains occurred in longer exposure to the stress condition as was found under high temperature and low soil moisture conditions (Takao et al., 1996). This finding therefore was in support to state that these roots were also under anatomical malformation. Soprano and Koller (1996) suggested that plant growth in citrus was restricted due to unavailability of trace elements at higher pH. In the present study, root injuries were distinct in the trees under pH 6.0-6.3 and 7.5-7.6. The degradation of starch grains, in addition, indicated the slowing of metabolic activities of plants. Therefore, this report is probably indicating the mechanism of restriction or unavailability of trace elements through root injuries. The degradation of starch grains close to the injurious sites may be an adaptive mechanism to compensate and endure the adverse condition causing injuries of roots. Naturally if the stored product is utilized for enduring the adverse environment, the normal growth of plants can not be expected.
The changing of root tip shape from pointed to dome and secretion of mucilaginous substances in the root tip under pH 7.5-7.6 indicated that root growth was also under cessation process which was more conspicuously evident by the shortening of meristematic region. Similarly Mohammad et al. (1996) found changes of root tip shape from pointed to a dome under high temperature and low soil moisture condition, Epidermal and root tip injuries under pH 7.5-7.6 similarly indicated the stern effects of higher culture solution acidity. The complete peeling of epidermal cells leaving the cortex bare may be due to the long exposure of roots under this condition. Due to injuries in the root tip, the growing region was also destroyed. This injury therefore suggested the total growth cessation of root under this condition. The complete degradation of standard grains can also be regarded as the sign of exhausted roots which may be their pre-death state.
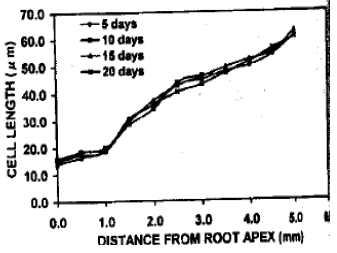 | |
| Fig. 5: | The cell length at different distances from the ap of roots of trifoliate orange budded with satsuma mandarin under pH 4.5-5.6 |
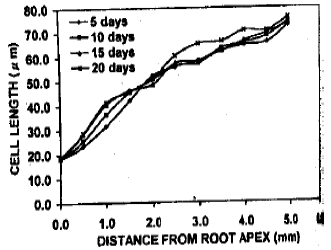 | |
| Fig. 6: | The cell length at different distances from the of roots of trifoliate orange budded with satsuma mandarin under pH 6.0-6.3 |
To understand the functional significance of the cell length profile, contrasting demands of intercellular and intracelluer transport should be considered. Intracellular transport is greatly facilitated by small cell size, while intercellular transport is impeded by the presence of cell wall. Developing cells are metabolically very active, so that in developing tissue a small cell size is necessary for the elevated rates of protein production. in contrast, intercellular transport is facilitated by longer cells. The meristem must import sucrose and other metabolites from the phloem and phloem is functional many cell lengths from the meristem. Although plasmodesmatal frequency and structure may be modified to facilitate sympiasmic transport, imports of metabolites is impeded to some extent by the cell wall. In stressed tissue, movements of metabolites from the phloem into the meristem is facilitated by two anatomical adaptations. Slow growth is usually associated with phloem differentiation close to the meristem (Rost and Baum, 1988). The increase in cell length at the base of the meristem also promotes intercellular transport. The comparatively larger and rounded cells near the root apex under higher pH therefore indicated these conditions as adverse and the elongated cells towards the root base under pH 4.5-5.6 stood in favor to opine the condition as ideal in our study.
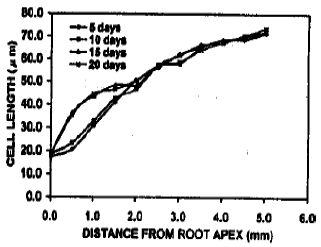 | |
| Fig. 7: | The cell length at different distances from the apex of roots of trifoliate orange budded with satsuma mandarin under pH 7.5-7.6. |
In conclusion, the suitable pH range was considered since a long ago as 6.0 to 6.5 for citrus. However, our study finds that the pH range 4.5-5.6 is better than that of 6.0 with respect to anatomical development which is known to be related with metabolic activities (Erickson, 1968). Our ultrastructural studies on this issue are going on. It can be expected that those studies on different cell organelles will facilitate us to show the consequences of culture solution acidity in the molecular level of roots of trifoliate orange budded with satsuma mandarin.
ACKNOWLEDGMENTS
We would like to thank Mr. A. lefuji and E. K. Tsuzuki of Citriculture Laboratory, Faculty of Agriculture, Ehime University, Japan for their partial assistance in this study.
REFERENCES
- Bertaud, D.S. and P.W. Gandar, 1985. Referential descriptions of cell proliferation in roots illustrated using Phleum pratense L. Int. J. Plant Sci., 146: 275-287.
CrossRefDirect Link - Brunet, J., 1994. Interacting effects of pH, aluminium and base cations on growth and mineral composition of the woodland grasses Bromus benekenii and Hordelymus europaeus. Plant Soil, 161: 157-166.
CrossRefDirect Link - Feigin, A., C. Ginzburg, S. Gilead and A. Ackerman, 1986. Effect of NH4/NO3 ratio in nutrient solution on growth and yield of greenhouse roses. Acta Hortic., 189: 127-136.
CrossRefDirect Link - Koyama, H., T. Toda, S. Yokota, Z. Deqair and T. Hara, 1995. Effects of aluminum and pH on root growth and cell viability in Arabidopsis thaliana strain Landsberg in hydroponic culture. Plant Cell Physiol., 36: 201-205.
CrossRefDirect Link - Medappa, K.C. and M.N. Dana, 1970. The influence of pH, CA, P and FE on the growth and composition of the cranberry plant. Soil Sci., 109: 250-253.
Direct Link - Takao, K., S. Masaya and P. Mohammad, 1996. Effects of high temperature and dry condition of summer season on the changes of root tip morphology in satsuma mandarin (Citrus unshiu Marc. cv. Okitsu Wase). Proceedings of the Symposium and Spring Meeting of Japanese Society for Horticultural Science, Volume 65, (JSHS’96), Japan, pp: 60-61.
Direct Link - Mohammad, P. and M. Shiraishi, 1998. Anatomical observations on the accumulation and utilization of storage starch and epidermis-related development in roots of vigorous and non-vigorous satsuma mandarin trees from early winter to early summer. J. Jap. Soc. Hortic. Sci., 67: 660-670.
CrossRefDirect Link - Mohammad, P. and M. Shiraishi, 1999. The effects of culture solution pH on root formation, root growth and root morphology in trifoliate orange budded with satsuma mandarin. Pak. J. Biol. Sci., 2: 14-20.
CrossRefDirect Link - Nonami, H. and J.S. Boyer, 1989. Turgor and growth at low water potentials. Plant Physiol., 89: 798-804.
Direct Link - Pritchard, J., R.G.W. Jones and A.D. Tomos, 1991. Turgor, growth and rheological gradients of wheat roots following osmotic stress. J. Exp. Bot., 42: 1043-1049.
CrossRefDirect Link - Rosen, C.J., D.L. Allan and J.J. Luby, 1990. Nitrogen form and solution pH influence growth and nutrition of two Vaccinium clones. J. Am. Soc. Hortic. Sci., 115: 83-89.
Direct Link - Rost, T.L. and S. Baum, 1988. On the correlation of primary root length, meristem size and protoxylem tracheary element position in pea seedlings. Am. J. Bot., 75: 414-424.
Direct Link - Silk, W.K., E.M. Lord and K.J. Eckard, 1989. Growth patterns inferred from anatomical records empirical tests using longisections of roots of Zea mays L. Plant Physiol., 90: 708-713.
Direct Link - Soprano, E. and O.L. Koller, 1996. Effect of lime incorporation depth and lime levels on the development of two citrus rootstocks under greenhouse conditions. Proceedings of the 8th International Citrus Congress, Sun City, South Africa, Volume 2, (ICC'96), International Society of Citriculture, Nelspruit, pp: 832-835.
- Yokomizo, H. and M. Ishihara, 1973. Studies on the mineral nutrition of fruit trees in sand and water culture. I. Effects of nutrient solution composition on the growth of satsuma trees. Bull. Hortic. Res. Station Series A, 12: 29-77.
Direct Link - Zieslin, N. and P. Snir, 1989. Responses of rose plants cultivar Sonia and Rosa indica major to changes in pH and aeration of the root environment in hydroponic culture. Scient. Hortic., 37: 339-349.
CrossRefDirect Link - Zieslin, N. and M. Abolitz, 1994. Leakage of phenolic compounds from plant roots: Effects of pH, Ca2+ and NaCl. Scient. Hortic., 58: 303-314.
CrossRefDirect Link