
S. E. Aguja
The United Graduate School of Agricultural Sciences, *Citriculture Laboratory, Faculty of Agriculture, Ehime University, 3-5-7 Tarumi, Matsuyama 790-8566, Japan
P. Mohammad
The United Graduate School of Agricultural Sciences, *Citriculture Laboratory, Faculty of Agriculture, Ehime University, 3-5-7 Tarumi, Matsuyama 790-8566, Japan
M. Shiraishi
The United Graduate School of Agricultural Sciences, *Citriculture Laboratory, Faculty of Agriculture, Ehime University, 3-5-7 Tarumi, Matsuyama 790-8566, Japan
T. Soga
The United Graduate School of Agricultural Sciences, *Citriculture Laboratory, Faculty of Agriculture, Ehime University, 3-5-7 Tarumi, Matsuyama 790-8566, Japan
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 2 | Page No.: 571-574







ABSTRACT
The ontogeny of chloroplast in satsuma mandarin (Citrus unshiu Marc. cv. Okitsu Wase) young leaves sprayed with urea was studied under transmission electron microscope. Leaves at 20 days after budding (DAB) exhibited undifferentiated mesophyll cells containing chloroplasts at different developmental stages for both control and urea-applied trees. However, in urea-applied trees the chloroplasts at different developmental stages were usually smaller and devoid of starch grains. Both control and urea-applied trees had almost similar pattern of chloroplast development which yielded six stages and are described in this paper.
PDF Abstract XML References Citation
How to cite this article
S. E. Aguja, P. Mohammad, M. Shiraishi and T. Soga, 1999. Ontogeny of Chloroplast in Satsuma Mandarin Young Leaves Sprayed with Urea. Pakistan Journal of Biological Sciences, 2: 571-574.
DOI: 10.3923/pjbs.1999.571.574
URL: https://scialert.net/abstract/?doi=pjbs.1999.571.574
DOI: 10.3923/pjbs.1999.571.574
URL: https://scialert.net/abstract/?doi=pjbs.1999.571.574
INTRODUCTION
Plastids are derived from the proplastids in the meristematic tissues (Muhlethaler, 1971; Pyke and Page, 1998) which differentiate into various types concurrent with tissue differentiation (Miyake and Maeda, 1976). Among the various types of plastids, chloroplasts are rather popular as these organelles are important sites of photosynthetic activities.
The control of plastid differentiation focused on the transition of proplastid to chloroplast development has been reported by Reiter et al. (1994) in Arabidopsis developing leaves as a model plant. Similarly, studies on the structure of plastids in some plant species other than citrus were also conducted (Miyake and Maeda, 1976; Williams, 1974; Dave and Rao, 1982; Berrie and Webster, 1982). Assuming that the structure of chloroplast vary from species to species or even within species among different tissues (Robertson and Laetsch, 1974; Kirchanski, 1975; Possingham, 1980; Thomson and Whatley, 1980), a thorough investigation of this organelle in a desired species is imperative. In citrus, ultrastructural studies particularly those dealing with plastic ultrastructure has been very limited.
For a more comprehensive information on the various functions of chloroplast in relation to its developmental stages, description of the different developmental stages is seemed necessary. Likewise, consequential effects of the external factors such as foliar nitrogen application in relation to chloroplast developmental stages needs also to be described. This could serve as an essential information
marking to further studies on chloroplast ultrastructure in nitrogen sufficient and deficient plants thus, the application of urea in this study has been considered.
ln this context, the present study was conducted in order to describe the different developmental stages of chloroplast in control and urea-applied satsuma mandarin young leaves.
MATERIALS AND METHODS
The experiment was conducted in the Citriculture Laboratory, Faculty of Agriculture, Ehime University, Japan during spring 1998. Three-year-old satsuma mandarin (Citrus unshiu Marc. cv. Okitsu Wase) trees of similar vigor were planted in potted sandy soil mixed with granite and maintained in plastic house with light intensity of approximately 60,000 lux and temperature range of 23-26°C. Heavy pruning was done by removing the old shoots and leaves on April 14, 1998. Spring flushes were allowed to grow for about 3-5 cm before spraying started. Three sprayings of urea (Wako Pure Chemicals Industries Ltd. Osaka, Japan) at 2,000 ppm were applied to 3 potted trees at 33ml/tree in the morning of April 28, 30 and May 2. The same number of trees were sprayed with tapwater which served as the control. Pots were covered with polyethylene bags to prevent entry of spray solution into the soil. Leaf sampling for 20 days after budding (20 DAB) was done on May 2, a day after the last spray.
The central portion of the leaf blades were cut into small pieces (1 mm x 2 mm) and were fixed in paraformaldehyde-glutaraldehyde for 24 hours at 10°C, postfixed in 1 percent osmium tetroxide for 2 hours at room temperature, dehydrated in a graded alcohol series and embedded in epoxy resin. Ultrathin sections were cut with diamond knife using the ultramicrotome (UT - 1000) and double stained with uranyl acetate and lead citrate. Sections were examined and photographed under HITACHI H-7100 transmission electron microscope at 100 kV.
A total of 130 photomicrographs (x 10,000) taken were considered in determining the different chloroplast stages for both urea-applied and control trees. Approximately 10 photomicrographs per developmental stage were used for measuring the chloroplast size on a length and width basis, and their averages were obtained.
RESULTS
Table 1 represents the size of chloroplast at different developmental stages in both urea-applied and control satsuma mandarin leaves at 20 DAB. The size of chloroplast increased as it developed, however, control trees had bigger chloroplasts. its size was about 30 percent larger than those of urea-applied trees.
| Table 1: | Size of chloroplast at different developmental stages in the mesophyll cells of urea-applied and control satsuma mandarin leaves 20 days after budding |
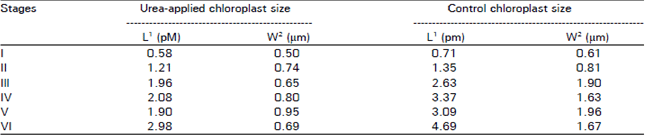 | |
| 1: Length; 2: width. Length and width were measured directly from the photomicrographs (x 10,000) | |
The different developmental stages of chloroplast in the mesophyll of urea-applied trees are illustrated in Fig. 1. The first stage of chloroplast development was considered as a plastid initial characterized by merely a small chloroplast with no internal structures (Fig. 1A). The second stage was figured out as proplastid invaginating internal membrane structure which showed infolding of membrane towards the inner portion of the chloroplast (Fig. 1B). The third stage involved internal lamellar structure formation marking the appearance of several thylakoids spreading to the sides perpendicular to the long axis of the chloroplast (Fig. 1C). The fourth stage exhibited an apparent formation of grana and the initial appearance of lipid droplets (Fig. 1D). The fifth stage entailed an extensive formation of stroma lamellar system exposing denser and longer thylakoids in the chloroplast (Fig. 1E). The sixth stage represented a full grown chloroplast characterized by the presence of developed grana, stroma thylakoids and several lipid droplets (Fig. 1F).
Fig. 2 represents the different developmental stages of chloroplast in control trees. Stage I shows a plastid initial which contained no internal structures similar to urea-applied treatment but bigger in size (Fig. 2A). Stage II is described as chloroplast containing initial internal structure characterized by the appearance of starch grains that almost filled the chloroplast (Fig. 2B). Stage Ill is featured by an internal lamellar structure formation around the strach grains (Fig. 2C). Chloroplast in stage IV is described where grana formation occurs around the starch grains and lipid droplets become apparent (Fig. 2D). Stage V is recognized with extensive stroma lamellar formation densely located in the central portion of the chloroplast and between starch grains (Fig. 2E). Stage VI is implicated as a full grown chloroplast filled with developed grana structures, stroma thylakoids, lipid droplets and starch grains (Fig. 2F). Lipid droplets in this treatment were lesser compared to urea-applied one (Figs: 1E, 1F, 2E and 2F).
Generally, the chloroplast ontogeny represented by different stages in both urea-applied and control treatments demonstrated a similar pattern, only that the chloroplasts in the control trees were bigger and contained starch grains.
DISCUSSION
Mesophyll cells of satsuma mandarin leaves at 20 DAB were still undifferentiated and this observation has been repeatedly shown in the previous studies (Aguja et al., 1996; Aguja and Shiraishi, 1998). However, at this leaf growth stage, internal organ differentiation particularly the plastid has already occurred, especially when the leaf nitrogen level is increased (Aguja and Shiraishi, 1998). Obviously, the smaller chloroplasts in urea-applied trees can be viewed as developing plastids which resulted from an active division of preexisting chloroplasts triggered by nitrogen application (Possingham, 1980). Whereas, the bigger chloroplasts in control trees can be considered as the same chloroplasts that preexisted in the cell and were not actively dividing but rather continued to develop. Likewise, Thomson and Whatley (1980) reported that during the early stages of cellular differentiation, variations in plastid size become apparently established as a result of differential expansion of the palisade prior to division. Moreover, as some cells become fully differentiated, plastids undergo dedifferentiation. This phenomenon is considerably suspected to occur in the chloroplasts of urea-applied trees where actively dividing chloroplasts take place triggered by the nitrogen application.
The different developmental stages of chloroplasts followed an almost similar pattern for both urea-applied and control trees. Except at the first stage in control trees, starch grains were always present in the chloroplast. This can be inferred that starch grain assimilation is slow in this treatment. Degradative enzymes for starch in these chloroplasts may be lacking due to low nitrogen content. Similar observations were reported in different nitrogen-deficient plant species exhibiting large starch grains in the chloroplasts (Thomson and Weier, 1962; Whatley, 1971; Chonan et al., 1977), The sequential pattern of chloroplast development particularly in urea-applied treatment was similar to that of Muhlethaler and Frey-Wyssling (1959). However, in control trees, invagination in the chloroplast was not observed at the second stage. Instead, an initial internal structure ina form of starch grain was seen. This difference in developmental plastid observation may be attributed to the vast occupation of starch grain in the chloroplast which may have accounted for the non-observation of invaginating (membrane. It may happen that in nitrogen-deficient plants this invagination stage is not clearly shown as starch grains may interfere with the process.
 | |
| Fig. 1: | Chloroplast developmental stages in urea-applied satsuma mandarin leaves 20 days after budding. A: Stage I - plastid initial; B: Stage II - invagination of internal membrane structure; C: Stage III-formation of internal membrane structure; D: Stage IV grana formation; E: Stage V - Stroma lamellae formation; F: Stage VI - full grown chloroplast. Bar 0.33 μm |
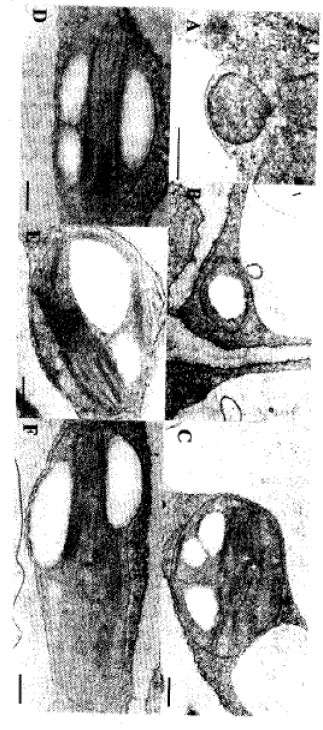 | |
| Fig. 2: | Chloroplast developmental stages in control satsuma mandarin leaves 20 days after budding. A: Stage I - plastid initial; B: Stage II - initial internal structure (appearance of starch grain); C: Stage III - formation of internal membrane structure; D: Stage IV - grana formation; E: Stage V - stroma lamellae formation; F: Stage VI - full grown chloroplast. Different chloroplast stages were contained with starch grains except for the first stage. Bar = 0.47 μm |
Moreover, this particular stage could be incorporated in the preceding and following stages of plastid development in nitrogen-deficient plants but can not only be seen clearly due to the presence of large starch grains. Certainly, in the event of plastid development, invagination really takes place.
In conclusion, the ontogeny of chloroplast in satsuma mandarin young leaves which yielded six stages was not altered by the application of urea. However, the ultrastructure of chloroplasts in urea-applied trees was different to the control in terms of size and starch grain content which were smaller and devoid of starch grain. These different developmental stages of chloroplasts for both urea-applied and control trees are described as follows: Stage I - a chloroplast considered as a plastid initial less than a micron in size and showing no internal membrane structures. Stage II - a chloroplast considered as a proplastid with invaginating internal membrane structure in urea-applied trees while in control, initial internal structure is formed represented by starch grain. Stage III - a chloroplast which involved formation of internal lamellar structures marking the appearance of several thylakoids spreading to the sides perpendicular to the long axis of the chloroplast. Stage IV - this stage is described as grana forming chloroplast coincided with the first appearance of lipid droplets. Stage V - a chloroplast representing a formation of stroma lamellar system exposing a denser and longer thylakoids. Stage VI - a stage recognized as a full grown chloroplast characterized by the presence of developed grana, stroma thylakoids and several lipid droplets.
REFERENCES
- Aguja, S.E. and M. Shiraishi, 1998. Ultrastructure of chloroplast in the young leaves of Satsuma mandarin top sprayed with different fertilizers under plastic house conditions. Asia Life Sci., 7: 109-119.
Direct Link - Berrie, G.K. and P.M. Webster, 1982. Ultrastructure of plastids and mitochondria in Gemmae of Marchantia polymorpha L. Ann. Bot., 50: 199-206.
CrossRefDirect Link - Chonan, N., H. Kawahara and T. Matsuda, 1977. Effect of nitrogen application on ultrastructure of the chloroplasts in rice plants. Jpn. J. Crop Sci., 46: 387-392.
CrossRefDirect Link - Dave, Y.S. and K.S. Rao, 1982. Plastid ultrastructure in the cambium of teak (Tectona grandis L. f.). Ann. Bot., 49: 425-427.
Direct Link - Kirchanski, S.J., 1975. The ultrastructural development of the dimorphic plastids of Zea mys L. Am. J. Bot., 62: 695-705.
Direct Link - Miyake, H. and E. Maeda, 1976. The fine structure of plastids in various tissues in the leaf blade of rice. Ann. Bot., 40: 1131-1138.
CrossRefDirect Link - Possingham, J.V., 1980. Plastid replication and development in the Life cycle of higher plants. Ann. Rev. Plant Physiol., 31: 113-129.
CrossRefDirect Link - Pyke, K.A. and A.M. Page, 1998. Plastid ontogeny during petal development in Arabidopsis. Plant Physiol., 116: 797-803.
Direct Link - Reiter, R. S., S.A. Coomber, T.M. Bourett, G.E. Bartley and P.A. Scolnik, 1994. Control of leaf and chloroplast development by the Arabidopsis gene Pale cress. Plant Cell, 6: 1253-1264.
Direct Link - Robertson, D. and W.M. Laetsch, 1974. Structure and function of developing barley plastids. Plant Physiol., 54: 148-159.
Direct Link - Thomson, W.W. and T.E. Weier, 1962. The fine structure of chloroplasts from mineral-deficient leaves of Phaseolus vulgaris. Am. J. Bot., 49: 1047-1055.
Direct Link - Thomson, W.W. and J.M. Whatley, 1980. Development of nongreen plastids. Ann. Rev. Plant Physiol., 31: 375-394.
CrossRefDirect Link - Whatley, J.M., 1971. Ultrastructural changes in chloroplasts of Phaseolus vulgaris during development under conditions of nutrient deficiency. New Phytol., 70: 725-742.
CrossRefDirect Link - Williams, E., 1974. Fine structure of vascular and epidermal plastids of the mature maize leaf. Protoplasma, 79: 395-400.
CrossRefDirect Link