
M. A. Elwakil
Plant Pathology Department, Faculty of Agriculture, Mansoura, University, Egypt
K. M. Ghoneem
Plant Pathology Department, Faculty of Agriculture, Mansoura, University, Egypt
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 1 | Page No.: 38-44







ABSTRACT
Eighty-six samples of blonde psyllium seed were collected from commercial markets in Egypt and tested for seed borne fungi. Both blotter and deep freezing methods were used to detect the maximum number of internal and external seed-borne fungi. A total of 41 species of fungi belonging to 21 genera were identified using the two selective methods. The pathogenicity of Fusarium oxysporum, F. solani and F. moniliforme was studied in psyllium seeds and seedlings. Both seed and soil inoculation were used in the studies. A higher percentage of pre- and post-emergence damping off developed from seed inoculation with each of three fungi than from soil infestation. The transmission of Fusarium oxysporum, F. solani and F. moniliforme from seed to mature plants of blonde psyllium was studied. All three fungi were found in root, hypocotyl and lower stem tissues of psyllium plants at intervals of 35, 60 and 120 days post-germination.
PDF Abstract XML References Citation
How to cite this article
M. A. Elwakil and K. M. Ghoneem, 1999. Detection and location of Seed-Borne Fungi of Blonde Psyllium and their Transmission in Seedlings. Pakistan Journal of Biological Sciences, 2: 38-44.
DOI: 10.3923/pjbs.1999.38.44
URL: https://scialert.net/abstract/?doi=pjbs.1999.38.44
DOI: 10.3923/pjbs.1999.38.44
URL: https://scialert.net/abstract/?doi=pjbs.1999.38.44
INTRODUCTION
Three species of the family Plantaginaceae, Plantago psyllium (P. aftra), P. indica and P. ovata, under the name of Plantago, have pharmaceutical uses to treat dysentery, chronic constipation and chronic diarrhea and as laxatives, demulcents emollients and diuretics (Wallis, 1967, Trease and Evans, 1978, Hussein, 1985, Chevallier, 1996, Nordgaard et al., 1996 and Chiej, 1988). According to the British Pharmaceutical Codex, the first two species are included as psyllium, while Plantago ovata is the source of seed husk. Plantago ovata was shown to produce a higher level of mucilage than the other two species and was chosen for the present investigation.
In compiling the list of seed-borne fungi attacking psyllium no attempt has been made to discriminate between seed infection, in which infected seed produces infected plants or is the focus of an epidemic and seed transport, in which an organism is present on the seed but has not been shown to subsequently cause infection.
The following fungi have been shown to affect the different species of Plantago (Kuiper and Bos, 1992).
Alexander (1982) and Alexander et al. (1984) studied the impact of an inflorescence disease caused by Fusarium moniliforme var. subglutinans on P. lanceolata. One of its effect was a reduction in seed production. De Nooij et al. (1987) studied an ear stalk disease on Plantago lanceolata caused by Phomopsis subordinaria (Desm.) Trav. Weevils transport the fungus into the stalk in or just below the ear (De Nooij, 1988 and Linders et al., 1996).
The impact of soil-borne pathogens on seeds, seedling and vegetative growth is of minor importance. Two fungal species, Fusarium oxysporum and Gliocladium roseum, could be regularly islotaed from moldy P. lanceolata seeds in grassland soil (Kuiper and Bos, 1992). This was regarded as saprophytic colonization on dead seeds.
Plantago wilt, caused by Fusarium oxysporum, was first reported in the United States (Russell, 1975). The same pathogen and F. solani were also reported in India (Naresh-Mehta, et al., 1985). In the annotated list of seed-borne pathogens of Richardson (1990), only two fungi, Peronospora plantaginis and Pythium ultimum, are recorded as seed-borne on psyllium seeds. Patel et al. (1984) reported a leaf disease caused by Alternaria alternata on Plantago ovata in India. Under severs conditions, shriveling and blackening of the seeds was also noticed.
Since scanty information on the diseases of blonde psyllium is available, the present work was planned to:
| 1. | Detect, identify and survey the seed-borne fungi of local and introduced psyllium seed in Egypt. |
| 2. | Study the nature of fungi isolated from seed and their effects on the psyllium plant. |
| 3. | Elucidate the mode of transmission of some pathogenic fungi from seeds to mature plants. |
Materials and Methods
Collection of seed samples: Eighty-six samples of blonde psyllium seed, collected from commercial markets in different parts of Egypt (including Gharbia, Alexandria, Cairo, Assuit, Sohag, Damietta and Dakhlia) during the years of 1994 and 1995, were used in the present study.
Seed health testing: The detection of seed-borne mycoflora was carried out following the Rules of the International Seed Testing Association (ISTA, 1993). Two hundred seeds of each sample were tested using standard blotter and deep-freezing methods.
Blotter method: Twenty-five seeds were plated in a 9-cm diameter Petri-dish containing three layers of moist blotters. The plates were incubated at 20±2°C for 7 days under cool white fluorescent lights with alternating cycles of 12 hours light and 12 hours darkness.
Deep freezing method: After plating the seeds as described in the blotter method, the dishes were incubated at 20°C for 24 hours and transferred to -20°C in a freezer for 24 hours. This was then followed by 5 days incubation at 20±2°C under cool white fluorescent lights with alternating cycles of 12 hours light and 12 hours darkness. The incubated seeds were examined after 7 days under a stereoscopic microscope (6-50X magnification) for the presence of fungi and their characteristics. Whenever necessary, the compound microscope was used to conform the identification after having examined the morphology of conidia and conidiophores. The seed-borne fungi were identified using the Commonwealth Mycological Institute, Kew, Surrey, England (CMI) description sheets, Danish Government Institute of Seed Pathology (DGISP) publications and papers by Booth (1985), Burrges et al. (1988), Chidambaram et al. (1973), Ellis (1971), Raper and Fennel (1965) and Singh et al. (1991).
With the aid of the stereoscopic microscope, hyphal tips from the fungi were transferred to potato dextrose agar (PDA) plates using the tips of heat-stretched capillary tubes. Pure cultures of the fungi were obtained and all isolates were maintained on PDA slants.
Pathogenicity test of important seed-borne fungi: Three species of Fusarium, F. oxysporum, F. solani F. moniliforme, were tested for their pathogenic effects on blonde psyllium seeds and seedlings.
Preparation of inocula: Seed lots showing a high incidence of infection with the important seed-borne fungi, Fusarium oxysporum, F. solani F. moniliforme, were plated on blotters. A micromanipulator was used to isolate the spores of each fungus. Capillary tubes were stretched using a Bunsen burner and the very fine tips were used to isolate the spores of the fungus under a stereoscopic microscope. The isolated spores were transferred to PDA media and incubated for several days in the dark at 24±2°C. The incubation period different from one fungus to another and from one isolate to another of the same fungus. When hyphal growth on the PDA in the Petri-dish reached 3 cm in diameter, one 0.5 cm disk of the culture was transferred to 50 ml of potato dextrose broth media. This was incubated in the dark for several days at 24±2°C until the hyphal mat covered the open surface of the liquid medium. Pure cultures of each fungus were maintained on PDA slants for further studies. The hyphal mats were harvested, washed with sterile distilled water and 50 grams of fresh mat of each fungus was blended in 500 ml of sterile distilled water to produce a sticky suspension. The prepared suspensions were used for inoculating the seeds and soil.
Seed dressing with the isolated fungi: Blonde psyllium seeds of pathogen-free seed lots were disinfected in 70 percent ethyl alcohol for 5 min and placed on sterilized tissue paper at room temperature to dry. Alcohol was used for the disinfestation process because seed mucilage resulted when using other disinfectants. Sterile seeds were then soaked in the inoculum suspensions for 5 min and immediately sown in plastic bags containing sterilized soil.
Conventional growing-on test: Both inoculated and non-inoculated seeds were sown (5 seeds per bag) in plastic bags (8 cm diam.) containing autoclaved sandy soil and allowed to grow under ambient conditions during the winter season. For one month, daily observations for germination and infection symptoms of pre- and post-emergence damping off were made and recorded.
Soil infestation with pathogenic fungi: Fifty grams of fresh mat of each fungus was blended in 500 ml of sterile distilled water to produce a sticky suspension. The prepared suspensions were added to 2 kg of peatmoss and mixed well. About 150 grams of this peatmoss was added to each plastic pot (8 cm diam.) containing 350 grams of steam sterilized sandy soil and mixed thoroughly. Sixteen replicates (pots) were used per treatment. The pots were kept in the greenhouse for seven days (the temperature ranged from 20 to 24°C) to allow the fungus to spread through the soil before sowing seeds. Another 16 pots were prepared in the same manner but without the fungus and served as a check. During the period the soil was kept moist by watering when needed.
Disease-free seeds were planted (5 seeds/pot) in infested and non-infested soil and kept in a greenhouse during the winter season. For one month, daily observations for germination and infection symptoms of pre- and post-emergence damping off were recorded.
Transmission of certain pathogenic fungi from seed to mature plants: Psyllium seeds were artificially inoculated with Fusarium moniliforme, F. oxysporum and F. solani. The same methods of inoculum preparation, seed inoculation, soil sterilization and sowing in plastic bags were followed as were used for the pathogenicity test. Ten replicates of plastic bags containing seeds inoculated with each fungus were prepared with 6 seeds sown in each bag. The seedlings emerging from the inoculated seeds were rated and left to grow. The transmission rates of the fungi at intervals of 35, 60 and 120 days were followed.
At the end of each interval, 10 plants were removed from the bags, washed, disinfected and dissected under sterile conditions. The different plant parts (roots, hypocotyl, lower stem, upper stem, peduncle, inflorescence and seeds if present) for each treatment were plated on sterile moist blotters and incubated for 7-10 days at 24°C. Fungi recovered for each treatment were identified and the transmission percentage was recorded.
Results
Seed health testing: A total of 41 species of fungi belonging to 21 genera were identified i.e., Alternaria alternate (Fr.) Keissler, A. tenuissima (Kunze expers.) Witshire, A. chlamydospora Mouchacca, A. longissima Deighton and MacGarvie, A. tenuissima (Kunze:Fr.) Wiltshire, Aspergillus flavus Link:Fr., A. fumigatus Fres., A. niger van Tieghem, A. ochraceus Wilhelm, A. tamarii Kita, A. clavatus Desm., A. carneus Blochwitz, Botrytis cinerea Pers.:Fr., Cephalosporium sp., Cladosporium sp., Curvularia affinis Boedijn, lunata (Wakker) Boedijn, C. pallescens Boedijn, Drechslera australiensis (Bugni:) Subram.andJain ex. M.B. Ellis [ = Bipolaris australiensis (M.B. Ellis) Tsuda and Ueyama], D. halodes (Drechs.) Subram and Jain, D. hawallensis (Bugni:) Subram and Jain ex. M.B. Ellis [ = Bipolaris hawaiiensis (Ellis) Uchida and Aragaki], D. Iongirostrata (Subram.) Subram, D. microbe (Drechs.) Subram and Jain, D. rostrata (Drechs.) Richardson and Fraser [= Exserohilum rostratum (Drechsler) Leonard and Suggs emend. Leonard], D. tetramera (McKinney) Subram. and Jain, Epicoccum sp., Fusarium moniliforme Sheld. F. oxysporum Schlecht, F. semitectum Berk. and Ray. [= F. pallicloroseum (Cooke) Sacc.], F. solani (Mart.) Sacc., Macrophomina phaseolina (Tassi) Gold., Mucor sp., Myrothecium indicum Rama Rao, M. vernIcaria (Albertini and Schwein.) Ditmar: Fr., Nigrospora sp., Penicillium sp., Phoma sp., Phomopsis sp., Rhizopus sp., Stemphylium sp., Trichothecium sp. and Verticillium sp. The percentage of detected fungi for each species is presented in Table 1. In comparing the efficiency of the two methods, a higher number of fungi were recovered with the blotter method than with the deep-freezing method. Using the blotter method, the dominant fungi were Cladosporium sp. at a maximum rate of 74.4 percent and an infection range of 0.5-34 percent, Altemaria afternata at 72 percent and an infection range of 0.5-39.5 percent, Aspergillus flavus at 70.9 percent and an infection range of 0.5-95 percent and Stemphylium sp. at 60.5 percent and an infection range of 0.5-7.5 percent.
The deep-freezing method recovered the following fungi; Alternaria alternate at a maximum rate of 66.3 percent and an infection range of 0.5-37.5 percent, Aspergillus flavus at 64 percent and an infection range of 0.5-90 percent, Cladosporium sp. at 54.7 percent and an infection range of 0.5-40.5 percent and Stemphylium sp. at 52.3 percent and an infection range of 0.5-9 percent.
Table 1 shows that the blotter method was better for detecting Fusarium spp., including F. moniliforme at a maximum rate of 36 percent and an infection range of 0.5-10 percent, F. semitectum at 26.7 percent and an infection range of 0.5-3 percent and F. oxysporum at 5.8 percent and an infection range of 0.5-2 percent . The same fungi were observed with the deep-freezing method. The maximum rate of F. moniliforme reached 29 percent with an infection range of 0.5-3.5 percent, F. semitectum reached 12.8 percent with an infection range of 0.5-4.5 percent and F. solani reached 3.5 percent with an infection range of 1-1.5 percent . Only, one seed sample was found infected with F. oxysporum with 0.5 percent infection observed in the seeds.
The deep-freezing method was not effective for detecting Botrytis cinerea and Drechslera halodes. However, it did show a higher percentage for Curvularia pallescens, which reached 5.8 percent with an infection range of 0.5-4 percent.
Aspergillus flavus showed a high percentage of detection on psyllium seeds with both methods. It was also noted that most of the examined samples were infected with several saprophytic fungi. The percentages were generally low when the deep-freezing method was used in comparison with the blotter method. The deep freezing method was more suitable for detecting Penicillium sp., which reached 39.5 percent with an infection range of 1-4 percent in comparison with the blotter method, which reached 30.2 percent with an infection range of 1-5 percent.
Pathogenicity testing of seed-borne fungi: Observations of infection symptoms on seeds and seedlings were recorded 30 days after sowing. Pathogenicity test results are presented in Table 2 and 3. Symptoms were classified as either pre- or post-emergence damping off. Pre-emergence damping off consisted of non-germinated seeds covered with hyphal growth of the tested pathogen. Post emergence damping off consisted of seedlings with lesions in the crown area and typical dark thread-like infected roots. Wilt symptoms were also considered to be post-emergence damping off.
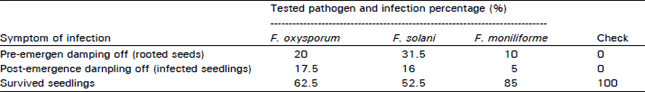
Effect of Fusarium oxysporum: Data presented in Table 2 and 3 show that the percentages of infection, expressed as symptoms of both pre-and post-emergence damping off, were 62.5 percent and 25 percent, respectively, for seeds and seedlings with the seed inoculation treatment, while they were 20 percent and 17.5 percent respectively, for seeds and seedlings from non-inoculated seeds planted one week after soil infestation with F. oxysporum. Percentage of surviving seedlings was higher (62%) with the soil infestation treatment than with seed inoculation (12%).
Effect of Fusarium solani: As shown in Table 2 and 3, the percentages of pre- and post-emergence damping off caused by Fusarium solani were 72.5 percent and 18.5 percent, respectively, for the seed inoculation treatment, while they were 31.5 percent and 52.5 percent, respectively, for the soil infestation treatment. The survival percentage of seedlings was 52.5 percent with soil infestation and 9 percent with inoculated seed sown in sterilized soil.
Effect of Fusarium moniliforme: The pre- and post- emergence damping off percentages caused by F. moniliforme were 15 percent and 13 percent, respectively, on seed and seedlings from inoculated seeds, while it was 10 percent and 5 percent, respectively, on sleds and seedlings from the soil infestation treatment.
| Table 1: | Fungi isolated from blonde psyllium seed samples |
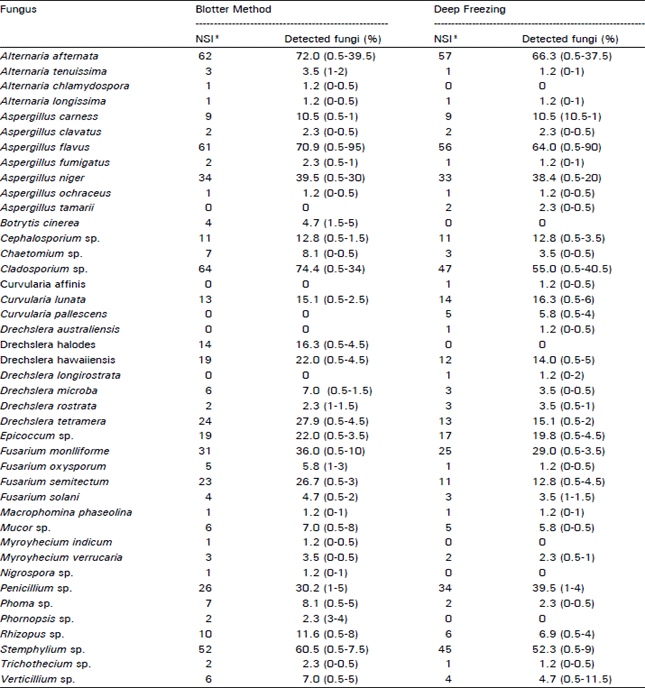 | |
| NSI: No. of samples infected (number in parentheses indicate infection range) percentage of detected fungi : (NSI / Total No. of samples (86)) X 100 | |
| Table 2: | Effect of Fusarium species (Fusasrium oxysporum, F. solani and F. monlliforme) on inoculated blonde psyllium seeds. |
 | |
| Table 3: | Effect of Fusarium species (Fusasrium oxysporum, F. solani and F. moniliforme) on inoculated blonde psyllium seeds and seedlings grown in infested soil. |
 | |
| Table 4: | Recovery percentage of fungi from seed to mature plants after artificial inoculation of psyllium seeds |
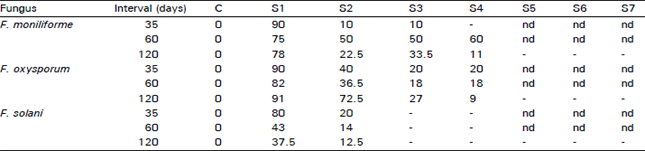 | |
| C = Check; S1 = Roots; S2 = Hypocotyl; S3 = Lower stem; S4 = Upper stem; S5 = Peduncle; S6 = Inflorescence; S7 Seeds; nd = not done due to the age of the plant. | |
Percentage of surviving seedlings was 85 percent in the soil infestation treatment compared with 72 percent in the seed inoculation treatment.
Data in Table 2 and 3 show that F. solani caused the highest infection rate of pre-emergence damping off in psyllium, followed by F. oxysporum and F. moniliforme. On the other hand, post-emergence damping off infection rates were the highest with F. oxysporum, followed by F. solani and F. moniliforme. Percentage of seedling survival was higher with F. moniliforme for both seed and soil infestation (being 72 percent and 85 percent, respectively) compared with the other two fungi.
Transmission of pathogenic fungi from seeds to mature plants: Among the fungi associated with psyllium seeds, three species of Fusarium, F. moniliforme, F. oxysporum and F. solani, were selected to study their transmission in a growing on test.
Table 4 shows that psyllium plants had different interactions with the fungi tested. All three tested fungi were isolated from roots, hypocotyls and lower stems of the plants at intervals of 35, 60 and 120 days. These intervals approximated the seedling, vegetative and reproductive plant growth stages. In the seedling stage (35 days from sowing), the percentage of recovery of F. moniliforme was 90 percent in the roots and 10 percent in the hypocotyl and lower stem. It was almost equal in all plant parts during the vegetative growth stage (60 days), with recovery from the roots at 75 percent and from the upper stem, 60 percent. In the mature plant (120 days), the fungus did not move beyond the upper stem.
Recovery of F. oxysporum from the stem showed that the fungus reached the top of the stem. Percentages of fungal recovery from the plated upper stem portions were 20, 18 and 9 percent after 35, 60 and 120 days, respectively. F. solani was the only fungus that appeared to be restricted to the roots and hypocatyls of the plants (infection rate was 12.5% at 120 days) and was not recovered in any of the aerial parts of the plants.
DISCUSSION
This study of the seed-borne fungi of psyllium seeds was undertaken because of their importance in the preparation of medicinal products. There was also a concern that some of the seed-borne fungi could pose a hazard to humans and animals due to mycotoxin production.
The various methods used in seed health testing differ in their sensitivity and purpose according to the nature of the seed and the pathogens associated with it. The blotter and deep freezing methods were selected in order to detect the maximum number of internal and external seed-borne fungi. The total number of fungal species detected on seeds was almost equal with the blotter (37 species) and deep freezing (36 species) methods. Six species, Altemaria chlamydospora, Botrytis cinerea, Drechslera halodes, Myrothecium indicum, Nigrospora sp. and Phomopsis sp., were exclusively found with the blotter method. Five species, Aspergillus tamarii, Curvularia affinis, C. pallescens, Drechslera australiensis and D. longirostrata, were only detected on the same seed samples when the deep freezing method was used.
Detection of the above-mentioned fungal species associated with psyllium seeds could be considered as the first comprehensive report on seed-borne fungi of blonde psyllium in Egypt. Our results agree in part with the findings of Russell (1975) and Mehta et al. (1985) who reported Fusarium oxysporum and F. solani on psyllium seed. A number of the fungi found to be seed-borne in blonde psyllium produce mycotoxins, i.e., Aspergillus flavus, A. ochraceus and Fusarium spp. This led us to suggest certain precautions, since the seeds are edible and are also used to produce pharmaceutical products. Public awareness is necessary before using the seeds. Seed-health testing should be done to avoid using potentially hazardous seeds. Little information regarding the seed-borne fungi of psyllium is available. Therefore, F. oxysporum, F. solani and F. moniliforme were selected to study their pathogenic effects on psyllium.
Data presented here clearly show variations among the different fungi in their pathogenic effects on germinability and seedling growth of psyllium seeds. Seed inoculation with F. oxysporum caused a higher percentage of seedling infection than soil infestation. Pre-emergence damping off was 62.5 percent with seed inoculation, while it was 20 percent with soil infestation. These results highlight the importance of seed infection in the production of the crop. The same trend was found with F. solani, which had 72.5 percent pre-emergence damping off with seed inoculation and 32 percent with soil infestation, while the post-emergence damping off was 19 percent and 16 percent, respectively. The survival of seedlings was 9 percent with seed inoculation and 53 percent with soil infestation. F. mondiforme was less severe, causing a moderate level of pre-emergence damping off (15 percent with inoculated seeds and 10 percent with seeds grown in infested soil, while it produced 13 percent and 5 percent post-emergence damping off, respectively).
Again, these data indicate the deleterious effect of seed-borne fungi on crop yields. The results agree with the findings of Dawar (1994) and Perveen (1996), who showed that artificially inoculated seeds gave a higher percentage of infection than artificially infested soil.
The above data on seed and soil infestation, which reflect the importance of seed-borne fungi rather than soil-borne ones, should remind the growers to use pathogen-free seeds and consider seed-health testing a must before planting. Moreover, using seeds of medicinal plants (e.g. blonde psyllium) which harbor mycotoxin-producing fungi could result in medical products contaminated with. mycotoxins.
It was found that the seed-borne fungi tested were able to infect seedlings and move up the stem to the inflorescence. Isolations showed that the fungi were moving upward through the vascular system of the stem. Only F. solani was restricted to the roots and hypocotyls of the plants.
ACKNOWLEDGMENTS
The authors are grateful to Dr. Conrad J. Krass, Primary State Plant Pathologist, State of California, Sacramento, CA, USA for his commentes on the manuscript and the improvement of the language.
REFERENCES
- Alexander, H.M., J. Antonovics and M.D. Rausher, 1984. Relationship of phenotypic and genetic variation in Plantago lanceolata to disease caused by Fusarium moniiforme var. subglutinans. Oecologia (Berlin), 65: 89-93.
Direct Link - Burrges, L.W., C.M. Liddell and B.A. Summerell, 1988. Laboratory Manual for Fusarium Research: Incorporating a Key and Descriptions of Common Species Found in Australia. 2nd Edn., Fusarium Research Laboratory, Department of Plant Pathology and Agricultural Entomology, University of Sydney, Sydney, Australia, ISBN-13: 9780949269560, Pages: 156.
- De Nooij, M.P., 1988. The role of weevils in the infection process of the fungus Phomopsis subordinaria in Plantago lanceolata. Oikos, 52: 51-58.
CrossRefDirect Link - De Nooij, M.P. and H.A. van der Aa, 1987. Phomopsis subordinaria and associated stalk disease in natural populations of Plantago lanceolate. Can. J. Bot., 65: 2318-2325.
CrossRefDirect Link - Linders, E.G.A., J.M.M. van Damme and J.C. Zadoks, 1996. Epidemics of Diaporthe adunca in experimental and in natural populations of Plantago lanceolata and the effect of partial resistance on disease development. Plant Pathol., 45: 70-83.
CrossRefDirect Link - Nordgaard, I., H. Hove, M.R. Clausen and P.B. Mortensen, 1996. Colonic production of butyrate in patients with previous colonic cancer during long-term treatment with dietary fiber (Plantago ovate Seeds). Scand. J. Gostroenterol., 31: 1011-1020.
CrossRefPubMedDirect Link