
Sahar Ali
Department of Biological Sciences, Quaid-I-Azam University, Islamabad, Pakistan
Asghari Bano
Department of Biological Sciences, Quaid-I-Azam University, Islamabad, Pakistan
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 4 | Page No.: 1305-1306







ABSTRACT
Despite that the number of nodules plant-1 and the diameter of pink bacteroid tissue were higher in Kinetin treatment as compared to other treatments, there was no significant effect determined on Acetylene reduction activity readings in different treatments. The four hormones IAA, ABA, GA and Kinetin did not show clear role on retarding or enhancing nodule senescence. There was no clear cut evident reflect the link between the senescence of upper portions of plant and nodule senescence.
PDF Abstract XML References Citation
How to cite this article
Sahar Ali and Asghari Bano, 1999. The Role of Growth Regulators on Nodule Senescence and Nitrogen Fixation in Soybean. Pakistan Journal of Biological Sciences, 2: 1305-1306.
DOI: 10.3923/pjbs.1999.1305.1306
URL: https://scialert.net/abstract/?doi=pjbs.1999.1305.1306
DOI: 10.3923/pjbs.1999.1305.1306
URL: https://scialert.net/abstract/?doi=pjbs.1999.1305.1306
INTRODUCTION
A very restrict symbiotic relation between soybean and rhaobium which lead to nodulation is determined by genotype of soybean root with Brady rhizobium jabonicum (Sadowsky and Kasslank, 1995). This symbiotic legume can enrich their soil (Thies et al., 1995). In soybean nature nodule were present in the pericycle cell in vascular bundle (Kouchi and Hata, 1993). The hormones which present in the nodules play an important role in early nodule development (Libbenga and Bogers, 1974). Nodule degeneration in legumes may occur naturally as a result of changes in environmental factors that affect plant growth or changes in plant growth (Sutton, 1983). The factors that can arrest nodule growth and development result in progressive deficiency in nitrogen fixing due to enhanced proteolytic activity (Atkins et al., 1984; Wagner and Sarath, 1987) and markedly inhibited shoot growth reduce in leaf protein level and chlorophyll break down (Atkins et al., 1984). Thomas and Stoddart (1975), pointed out factors which has been associated with the onset of senescence including genetics and/or hormones.
Seemingly that application of hormones particularly Kinetin should elongate the photosynthate to nodule and result in stimulated and prolonged nodule activity. To detect this bioactive different hormones were used.
Materials and Methods
Soybean seeds were obtained from Natural Agricultural Research Center (NARC), Islamabad. The seeds were soaked for 6 hrs. with aqueous solution of ABA, IAA, GA, Kinetin and distilled water each at 10-5M and 10-6M. The seeds were inoculated with Rhizobium Leguminosarum strain NARCI in 48 percent sugar solution. The seeds were sown in pots containing soil and organic matter (2:1) under natural condition and supplied by half strength Hoagland solution once a week.
Acetylene reduction activity was recorded after flowering stage following the method of LaRue and Kurz (1973). The number of nodules were determined and the volume of pink bacteroid tissue was determined by detecting the diameter of pink bacteroid tissue which is the responsible for N2 fixation in light microscope at 4X.
RESULTS AND DISCUSSION
A decline in N2 fixation after flowering is a characteristic of nodule senescence due to enhanced proteolytic activity (Wagner and Sarath, 1987). Since there was no significant different in the optical density value for ethylene converted from the acetylene by nodules in different treatments.
| Table 1: | Plant physico-chemical parameters as affected by growth regulators in Soybean |
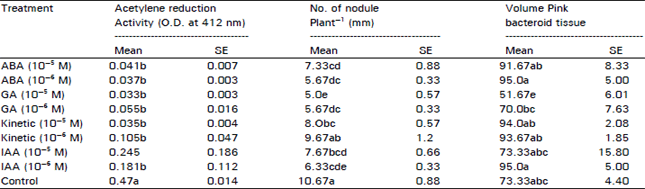 | |
The hormones have showed no significant effect on retarding or enhancing nodule senescence. Although the hormones had shown little effect on the treated plants presented in number of nodule plant‾1 and the volume of pink bacteroid tissue where Kinetin treated plant showed the higher records as compared to other treatments (Table 1).
On the other hand and under the concept of carbohydrate-nitrogen relationship which shed on the kind of relationship between nodule and upper portions of plant. Kinetin fail to retard nodule senescence on the time that Kinetin treated plants show delay in their leaf senescence. This can be explained by suggesting that depletion of carbohydrate during pod filling cause deficiency in carbohydrate which cause ammonia toxicity followed by nodule senescence (Sutton, 1983) and that mean some other factor rather than carbohydrate availability can terminate acetylene reduction activity (Malik, 1983). So clear link between leaf and nodule senescence did not overlooked in this experiment.
REFERENCES
- Atkins, C.A., B.J. Shelp, J. Kuo, M.B. Peoples and J.S. Pate, 1984. Nitrogen nutrition and the development and senescence of nodules on cowpea seedlings. Planta, 162: 316-326.
CrossRefDirect Link - Kouchi, H. and S. Hata, 1993. Isolation and characterization of novel nodulin cDNAs representing genes expressed at early stages of soybean nodule development. Mol. General Genet. MGG, 238: 106-119.
Direct Link - LaRue, T.A. and W.G.W. Kurz, 1973. Estimation of nitrogenase using a colorimetric determination for ethylene. Plant Physiol., 51: 1074-1075.
Direct Link - Malik, N.S.A., 1983. Grafting experiments on the nature of the decline in N2 fixation during fruit development in soybean. Physiol. Plantarum, 57: 561-564.
CrossRefDirect Link - Thies, J.E., P.W. Singleton and B.B. Bohlool, 1995. Phenology, growth and yield of field-grown soybean and bush bean as a function of varying modes of N nutrition. Soil Biol. Biochem., 27: 575-583.
CrossRefDirect Link - Thomas, H. and J.L. Stoddart, 1975. Separation of chlorophyll degradation from other senescence processes in leaves of a mutant genotype of meadow fescue (Festuca pratensis L.). Plant Physiol., 56: 438-441.
PubMedDirect Link