
Asghar Ali
Pulses Programme, National Agriculture Research Center, Park Road Islamabad, Pakistan
D.L. Johnson
Department of soil and crop sciences, Colorado state university, fort Collins, co 80523, USA
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 4 | Page No.: 1292-1295







ABSTRACT
To determine association between any two of the three traits of cold tolerance, growth habit and anthocyanin pigment in lentil (Lens culinaris Medikus), F2 populations were studied for two years (1991/92 and 1992/93) at Quetta and F3 for one year (1992/93) at two different field sites, Quetta and Kalat, Balochistan, Pakistan. Highly significant (p<0.001) association between any two of the three traits in all possible combinations was found which indicates that indirect selection for winter hardiness, using prostrate growth habit and anthocyanin pigment, is possible.
PDF Abstract XML References Citation
How to cite this article
Asghar Ali and D.L. Johnson, 1999. Association of Growth Habit and Anthocyanin Pigment with Winter Hardiness in Lentil. Pakistan Journal of Biological Sciences, 2: 1292-1295.
DOI: 10.3923/pjbs.1999.1292.1295
URL: https://scialert.net/abstract/?doi=pjbs.1999.1292.1295
DOI: 10.3923/pjbs.1999.1292.1295
URL: https://scialert.net/abstract/?doi=pjbs.1999.1292.1295
INTRODUCTION
Approximately 50 percent of the area of the Balochistan province of Pakistan is described as arid uplands (>1000 m) and is representative of the arid highlands of W. Asia (Ali et al., 1991). Highly variable and unpredictable environmental conditions (primarily cold and drought) prevail in the region where lentil is cultivated. With the improvement of winter hardiness acreage of lentil may be increased in the region which may lead to increase in the total production.
Direct selection for winter hardiness is the best approach for improvement of this trait in lentil but under natural conditions it is not possible without having differential winter kill in the field which can only be seen in certain number of years when the air temperatures are less than or equal to -15°C (personal observation). Furthermore, the natural environments are not reliable and controllable due to their various inherited problems. To speed up the breeding work and to improve its efficiency, it is essential to find an association among morphological traits for winter hardiness to make indirect selection for this trait in the breeding material of lentil when extreme low air temperatures in the field are not experienced.
There are few studies in other crop species indicate association among morphological traits for winter hardiness. The development of a compact rosette of shoots was found essential for winter survival in pea (Markarian and Andersen, 1966; Andersen and Markarian, 1968). Growth habit and cold tolerance have been found to be related in barley. The more upright types were injured more by freezing temperatures when compared to prostrate types under field conditions (Abo-Elenein et al., 1967). Reid (1965) established a definite association between winter growth habit and high winter survival but this association was not complete and hardy spring lines of barley were also isolated. Pea plants with high anthocyanin (purple pigment) have the greater number of cold hardy segregates (Markarian et al., 1968). However, the authors were not sure that the anthocyanin content itself was responsible for increased hardiness or if it was linked or associated with the factors responsible for winter hardiness. Ellis and Hong (1995) found that lentil genotypes which survive cold winters also have a lower cool base temperatures for germination, but their findings revealed that it is not possible to select cold tolerant genotypes directly from estimates of the base temperature for germination. The objective of this study was to determine association between any two of the three traits of cold tolerance, growth habit and anthocyanin pigment in lentil.
MATERIALS AND METHODS
Meteorological data were recorded in each year and at each of two field sites, Quetta (altitude 1750 m, latitude 30° 14' N, longitude 67° 2' E) and Kalat (altitude 1850 m, latitude 29° 07' N, longitude 66° 24' E), in Balochistan province of Pakistan. Above average rainfall (280 and 308 mm respectively) was received at Quetta in both growth seasons of 1991/92 and 1992/93. The minimum temperature in both seasons was -7°C. Although temperatures never fell below zero during January of the 1991/92 season but January, 1993 had 14 days below at Quetta. There were quite few genotypes injured during 1991/92. The low temperatures at the Kalat location were similar to Quetta. Substantially reduced rainfall (39 mm) during the growth period was received at Kalat.
Five different lines and/or cultivars, including winter hardy and spring lentils, were selected for crossing. All winter hardy lines had prostrate growth habit and anthocyanin pigment present whereas spring lentil had erect growth habit and pigment absent. Three males Balochistan Local, ILL 5865 (recently released variety in Pakistan as SHIRAZ-96) and ILL 5677 (all winter hardy) and two females, WA 8649090 (WA) (winter hardy) and Brewer (non-hardy spring lentil), were used in six one-way crosses following a factorial design (North Carolina Design-II) format (Comstock and Robinson, 1948). Two sets of F, seed were made. One set (F2 population) was planted at Quetta on October 19, 1991 and the second set on October 13, 1992 at the same site. At least six seeds were sampled from each F2 plant, bulked across all F2 plants within each cross and divided into two sets of two replications each to grow F3 bulk populations during the next season at Quetta and at Kalat. The experiments during the 1992/93 season were planted on October 12, 1992. In all experiments, all crosses were nested with their parents in families in two replications in the field. Each plot was space-planted with 140 seeds to obtain at least 128 plants per plot in a randomized complete block design (RCBD). The plant-to-plant and row-to-row distance was 50 cm. A pre-planting irrigation of c. 50 mm was applied to all plots to ensure emergence. The experiments were then dependent upon rainfall until, plant maturity. Soil at both sites was a sandy clay loam. Nitrogen and phosphate fertilizers at 20 and 60 kg ha‾1 respectively were incorporated into the soil at the time of seedbed preparation. Cold tolerance was scored on a scale of 1-5 where 1 = no foliar damage and 5= completely dead. It was recorded on March 27 when clear symptoms of cold injury after possible recovery of the plants could be seen (Auld et al., 1983; Ali et al., 1991). On 15th of December, when the plants were fully acclimatized, growth habit data were recorded on the scale of 1-3 where 1 was used for prostrate, 2 for semi-prostrate and 3 for erect growth habit. At the same time, pigment data were recorded as 1 = presence of pigment and 2 = absent.
A Statistical Analysis System (SAS) computer software program was used to analyse the data. Chi-square test of independence was used to determine the association between any two of the three traits of cold tolerance, growth habit and anthocyanin pigment (SAS, 1988).
RESULTS
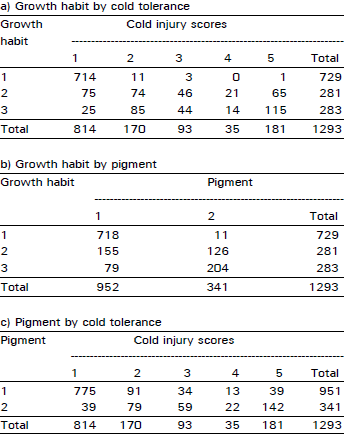
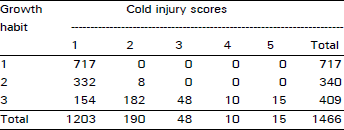
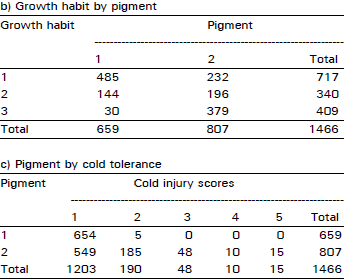
An extremely large number of genotypes having no cold injury also had prostrate growth habit and anthocyanin pigment present consistently in all plant populations in two years of field study across the locations and generations (Table 1-4). Using this data, chi-squared test of independence showed that all three traits (cold tolerance, growth habit and pigment) in all possible combinations in all experiments at all sites were highly significantly (p<0.00 1) associated (Table 5).
| Table 1: | Number of plants falling in each possible group in all possible combinations among the traits of cold tolerance, growth habit and anthocyanin pigment in F2 population at Quetta during 1991/92 crop season |
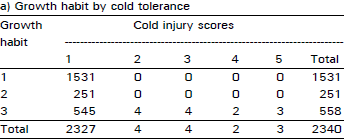 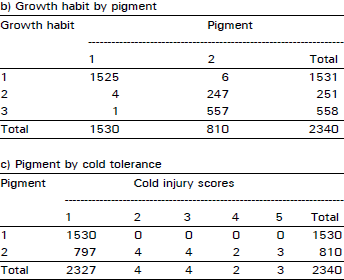 | |
| Table 2: | Number of plants falling in each possible group in all possible combinations among the traits of cold tolerance, growth habit and anthocyanin pigment in F2 population at Quetta during 1992/93 crop season |
 | |
DISCUSSION
Natural environments are highly inconsistent, unpredictable and non-repeatable. Moreover, confounding effected different factors like diseases, insect damage, frost heavy planting date and various other cultural practices encountered in the field make field selection for hardiness difficult and unreliable. Years/locations, such as in 1991/92 at Quetta, can make direct selection impossible and ultimately progress would be slow as the expression of cold tolerance is extremely dependent upon the severity of the environment. Indirect selection is a likely alternative in this kind of situation. A significant (p<0.001) association among cold tolerance, growth habit and pigmentation is evident, which may make selection for winter hardiness possible under field conditions when differential winter kill is not available.
| Table 3: | Number of plants falling in each possible group in all possible combinations among the traits of cold tolerance, growth habit and anthocyanin pigment in F3 population at Quetta during 1992/93 crop season |
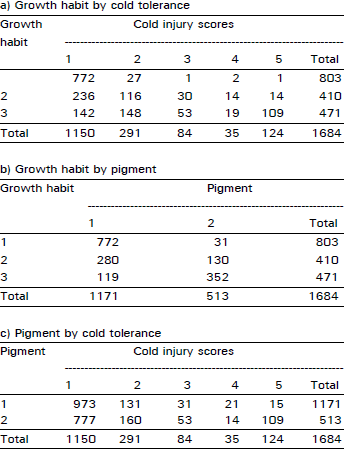 | |
| Table 4: | Number of plants falling in each possible group in all possible combinations among the traits of cold tolerance, growth habit and anthocyanin pigment in F3 population at Kalat during 1992/93 crop season |
  | |
| Table 5: | Chi-squared values for the test of independence of traits of growth habit (Gh), cold tolerance (Ct) and pigment (Pm) in all possible combinations in all populations, years and locations. All values are highly significant (p<0.001), n = sample size |
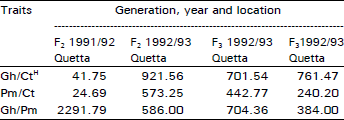 | |
Large numbers of plants having prostrate growth habit and pigment were not injured at all in the field during 1992/93 at both sites when damage to other plants in response to low temperatures was observed. This indicates that indirect selection for winter hardiness can be made by using prostrate growth habit and purple pigmentation.
ACKNOWLEDGMENTS
The authors acknowledge the financial support of USAID and PARC under MART/AZR project for Ph.D. programme and the research work reported in this paper. We gratefully acknowledge the seed provided by W. Erskine (ICARDA) and F. J. Muehlbauer, (WSW and the literature and helpful suggestions provided by J. D. H, Keatinge (University of Reading) during this course of study.
REFERENCES
- Ali, A., J.D.H. Keatinge, B.R. Khan and S. Ahmad, 1991. Germplasm evaluation of dual-season lentil (Lens culinaris) lines for the arid highlands of West Asia. J. Agric. Sci., 117: 347-353.
CrossRefDirect Link - Andersen, R.L. and D. Markarian, 1968. The inheritance of winter hardiness in Pisum. III. Stem branching in autumn growth. Euphytica, 17: 473-478.
CrossRefDirect Link - Auld, D.L., K.J. Adams, J.B. Swensen and G.A. Murray, 1983. Diallel analyses of winterhardiness in peas. Crop Sci., 23: 763-766.
CrossRef - Markarian, D. and R.L. Andersen, 1966. The inheritance of winter hardiness in Pisum I. Euphytica, 15: 102-110.
Direct Link - Markarian, D., R.R. Harwood and P.R. Rowe, 1968. The inheritance of winter hardiness in Pisum II. Description and release of advance generation breeding lines. Euphytica, 17: 110-113.
Direct Link - Reid, D.A., 1965. Relationship of growth habit, winter hardiness and qualitative genetic markers in barley (Hordeum vulgare L. Emend. Lam.). Crop Sci., 5: 557-561.
Direct Link