
Nagwa M. Ateef
Botany Department Faculty of Science, Cairo University Giza, Egypt
Nagwa M. Morsi
Botany Department Faculty of Science, Cairo University Giza, Egypt
Magdy El-Said Mohamed
Botany Department Faculty of Science, Cairo University Giza, Egypt
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 4 | Page No.: 1211-1218







ABSTRACT
The biodegradation of two light crude oils from lad and marine wells at Red Sea area by a bacteria enrichment culture was studied under different environmental factors. Enhanced oil biodegradation was observed with aeration or shaking, mesophilic temperature range and salinity between 3-7 percent. Preferential use of urea as nitrogen source resulted in 4-5 fold increase in oil consumption. Nitrogen/phosphorus molar ratios between 10:1 and 20:1 were optimal for oil degradation. Supplementation with iron stimulated biodegradation with optimal concentration of about 2.5 μ mol. Dramatic decrease in oil consumption was observed in absence of emulsifying agent. lnocula of increased optical densities resulted in increased oil consumption but not at the same magnitude. At optimal conditions, the bacterial culture consumed two thirds of the added oil in one week. Compositional variations with respect to C-, H-, N-, S-, P- contents and C/H ratio of the recovered oil were observed. Four bacterial genera naturally inhabiting the crude oil were isolated and identified as Micrococcus sp., Bacillus sp., Corynebacterium sp. and Pseudomonas sp.
PDF Abstract XML References Citation
How to cite this article
Nagwa M. Ateef, Nagwa M. Morsi and Magdy El-Said Mohamed, 1999. Crude Oil Biodegradation by Naturally Inhabiting Mixed Bacterial Culture under Different Environmental Factors. Pakistan Journal of Biological Sciences, 2: 1211-1218.
DOI: 10.3923/pjbs.1999.1211.1218
URL: https://scialert.net/abstract/?doi=pjbs.1999.1211.1218
DOI: 10.3923/pjbs.1999.1211.1218
URL: https://scialert.net/abstract/?doi=pjbs.1999.1211.1218
INTRODUCTION
The oil spill incidents and their severe persistent polluting impact on the environment is still a major world problem. Although the mechanical and chemical method used in the treatment of water and soil pollution resulting from oil spills (Aldous, 1991) have their powerful role the microbial tools still have their advantages over other methods. Although the microbial degradation of crude oil is a slow process, it is onset process that removes toxic components from the environment and helps in enriching the biogeochemical cycles of elements in the environment.
The ability of microorganisms to degrade a vast number of hydrocarbons of diverse structure present in petroleum has been addressed through many articles with different goals. Some studies have focused on the impact of oil spillage on water and soil-inhabitant microflora (Stafford et al., 1982; Barkay and Pritchard, 1988; Rahman et al., 1993). While other aimed at the production of single cell protein (Zaki and El-Badrawy, 1981; Zaki et al., 1982). Recently, researchers are concerned with illustration of the influence of biodegration on the chemical and physical properties of crude oils in many shallow reservoirs to permit better prediction of the nature of the recovered crude oils (Milner et al., 1977; Sassen, 1980; Wolcott et al., 1989). Biodegradation of petroleum in the environment is a complex process which is highly dependent on the environmental and nutritional conditions in addition to the physical and chemical state of the oil (Atlas and Bartha, 1972; Dibble and .Bartha, 1979; Atlas, 1981; Leahy and Colwell, 1990; Swannell et al., 1996). Light oils are more susceptible to biodegration than heavy oils depending on the type and percentage of the metabolizable fraction present in it (Leahy and Colwell, 1990; Walker et al., 1976). Conditions required to enhance oil biodegration in natural environment include presence of high oil/H20 interface, an influx of water bringing inorganic nutrients and dissolved oxygen, suitable temperature and well-adapted microbial culture. In addition, the role of essential inorganic nutrients, nitrogen, phosphorus and iron has been demonstrated (Atlas and Bartha, 1973; Dibble and Barth, 1976; Olivieri et al., 1976; Atlas 1981).
MATERIALS AND METHODS
Crude oils: Two light crude oils from producing land and marine wells at Red Sea area were used. Land oil is a 2.7 API crude with 2.3 percent sulfur and 0.005 percent salt, whereas, marine oil has gravity of 26.5 API, sulfur content of 2.5 and 0.015 percent salt (wt/wt).
Media: The minimal medium used, contained (g%): K2HPO4, 0.3; KH2PO4, 0.1; MgSO4. 7H20, 0.1 and Tween 20, 0.05; dissolved-in order-in distilled water. Nitrogen sources were added at 40 mM nitrogen in the form of urea, NaNO3, KNO3, NH4NO3, NH4CI, (NH4)2SO4 and (NH4)3PO4. Water-soluble iron was added as ferric ammonium citrate. The minimal medium was supplemented with about 2.8 g percent (wt/vol) crude oil as sole carbon and energy source. The pH of the medium was adjusted to 7.2. For salinity experiment, the medium described by Rodriguez-Valera et al. (1980) was used. Sterile air was forced through the medium at 4.51 air/min/250-ml flask to supply oxygen required for microbial growth.
Bacterial culture and incubation conditions: An oil-degrading bacterial culture isolated from marine oil sample was enriched and used in this study and was maintained on nutrient agar slants (Oxoid). Trials to isolate bacteria from land oil were not successful. The inoculum for each experiment was pregrown for 24 h on nutrient agar slants and collected by scratching in 3 ml of sterilized minimal medium lacking carbon and nitrogen sources. This method of inoculum preparation was followed to avoid the transfer of nitrogen and carbon sources from nutrient agar to the minimal medium. Each treated flask received 5 ml inoculum of optical density equal to one at 580 nm. The standard incubation conditions used, unless otherwise stated, mean one week incubation at 35°C, aeration at 4.51 air/min/250 ml flask and starting culture optical density of about 0.07 at 580 nm.
Bacterial culture enrichment: For enrichment of oil-degrading aerobic bacteria, sterile minimal medium lacking carbon source was supplemented with non-sterile oils int he ratio fo 1-3 g oil/100 ml medium in 250 ml Elenmeyer flask and kept at 30°C for two weeks under shaking at 120 rpm. The water phase of the medium acquired light grey colour which became opaque in case of marine oil. Microscopic examination of water phase and streaking on agar plates revealed the presence of mixed bacterial colonies. Gram-stained smear illustrated the presence of at least three to four morphologically distinct Gram-positive and negative strains. This bacterial enriched culture was maintained on nutrient agar.
Identification of bacterial isolates: Bacterial culture was separated from oil under aseptic conditions and purified using the streak method. The purified cultures were then subjected to different tests including microscopic analysis, staining procedures (Gram's stain and spore stain) in addition to biochemical and physiological analysis following the procedures adopted by Bergey's manual of determinative bacteriology (Buchanan and Gibbons, 1974; Mitruka and Bonner, 1976). Biochemical and physiological tests included catalase, oxidase, carobydrate fermentation, motility, oxygen requirements, fat hydrolysis, indole, gelatin liquefaction, starch hydrolysis, casein hydrolysis and urease.
Determinations: Nitrate, nitrite and ammonia were determined according to Benson (1994). Residual oil was recovered by two consecutive extractions using diethyl ether and determined gravimetrically after evaporation of solvent. Percentage of biodegradation was calculated by comparing the amount of residual oil to the amount of added oil after correction of the non-biological losses. Elemental analysis of oils was carried out in the Micro-Analytical Center (MAC), Faculty of Science, Cairo University. Bacterial biomass was collected by centrifugation of oil-free culture medium at 10,000 rpm for 10 min. All treatments and analysis were performed in duplicate. Analysis of variance by one way ANOVA (SPSS programme under windows, Microsoft, 1991, U.S. A.) was estimated for the obtained experimental data.
RESULTS
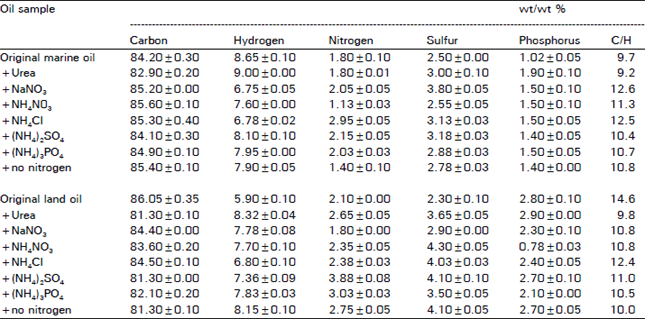
Crude oil analysis: The elemental analysis of both land and marine oils is given in Table 1. A marked difference in hydrogen and phosphorus contents as well as in C/H ratio of the two oils was observed. Both oils are considered as high-sulfur containing oils. The salt content of marine oil is three fold that of land oil.
Bacterial isolates: Purification ad staining procedures sowed the presence of four different bacterial isolates in marine oil. An isolate showed a Gram negative appearance and rod shaped, another isolate proved to be Gram positive cocci and two other Gram positive bacilli isolates. Following the used tests (materials and methods) the organisms were found to be Micrococcus sp., Bacillus sp., Corynebacterium sp. and Pseudomonas sp.
These isolates were used as a mixed bacterial culture in all the following experiments. More work to reach the species level of such genera is a further goal.
Biological and non-biological loss of oil: A marked decrease in the amount of recovered marine oil was observed. To ensure that the loss of marine oil is mainly due to the metabolic activities of the enriched bacterial culture, the scheme given in Table 2 was followed. Neither the set of flasks receiving sterilized oil was considered as non-biological loss.
The enriched bacterial culture isolated from the marine oil sample was employed in studying the influence of some environmental factors on oil biodegradation to establish the optimum conditions required to increase oil consumption.
Effect of different nitrogen sources: The different nitrogen sources were tested at 40 mmol nitrogen concentration (Table 3). The medium (100 mI/250 ml flask) supplemented with 2.8 g oil, 1 μM Fe and phosphate concentration was adjusted to give N/P molar concentration ratio fo 10:1. Flasks were inoculated with the previously isolated bacterial culture and kept at 35°C for 7 days. Sterile air was forced through it at 4.51 air/min/flask. Two sets of control were run in parallel, one set without exogenous nitrogen and the other was uninoculated. The pH value, NO3, NO2 and NH3 were measured through the incubation period. After 7 days incubation, weights of the residual oil as well as that of the microbial biomass in the water phase were determined.
The results showed that all tested nitrogen sources supporting biomass formation resulted in statistically significant stimulated oil consumption (Table 3). Urea was the best metabolized nitrogen source resulting in four to five fold increase in biodegradation of both oils over control lacking exogenous nitrogen. Nitrate as Na- or K-salts, NH4CI and NH4NO3 followed urea in enhancing oil biodegradation, whereas (NH4)2SO4 and (NH4)3PO4 were the least effective nitrogen sources. A pronounced decrease of residual oil accounting for about 6 percent (after subtracting the non-biological loss of oil) was observed in samples lacking external nitrogen source. In all cases, the nitrogen source supplemented to culture was detected in the medium throughout the whole biodegradation period (data not included). The cultures lacking external nitrogen source showed transient presence of nitrite.
| Table 1: | Elemental analysis and some physical properties of land and marine crude oils |
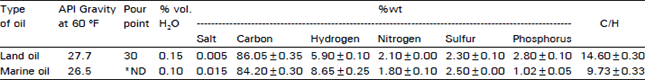 | |
| *ND: Not determined | |
| Table 2: | Scheme followed to determine the biological and non-biological loss of oil. The culture medium (100 ml) contained 2.5 g oil and was incubated at 30°C for two weeks under shaking (120 rpm) |
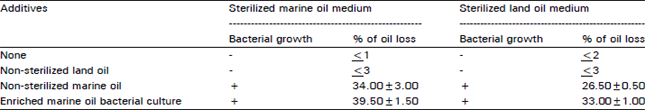 | |
| Table 3: | Effect of different nitrogen sources (40 mM N/l) on oil consumption and iomass formation in aerated cultures |
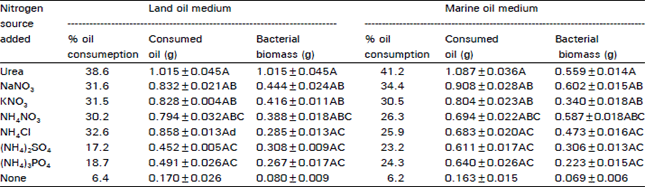 | |
Significance is calculated at 5 percent level.A: Significant in relation to control lacking exogenously added nitrogen B: Significant in relation to control as nitrate group. C: Signifidant in relation to control as amonium group | |
The results showed that consumption of both oils was nearly at the same rate. The pH in urea-containing medium was nearly. stable, while a slight change in pH was observed with other nitrogen sources containing media which ranged between 6.5 and 7.8.
From the pervious results, urea proved to be the most efficient nitrogen source in enhancing oil biodegradation.
Effect of static, shaked or aerated cultures on oil consumption: The isolated mixed bacterial cultures were inoculated into marine and land oil-containing media. The flasks were incubated under static, shaked (120 rpm) or aerated (0.9 102/min/250 ml flask) conditions. The utilization of some nitrogen sources in addition to urea were tested to reveal their Influence on oil consumption in relation to oxygen content of the culture (Fig. 1). The obtained results showed marked differences in the amount of consumed oil even with the same nitrogen source. Aerated cultures were the most potent oil consumers showing two to three fold increase in oil consumption over static ones. As previously noticed, urea proved to be superior over other nitrogen sources used in enhancing oil biodegradation under aerated, shaked or static cultures. In case of land-oil supplemented medium, the same results were obtained. Hence data concerning marine oil were only presented.
In the following experiments urea was used as the most efficient nitrogen source, while oil was superior over land oil under aerated conditions.
Effect of nitrogen/phosphorus ratio: Different concentrations of phosphorus were added to culture medium containing 40 mM nitrogen to establish nitrogen/ phosphorus molar ratios between 1:4 and 25:1 (Fig. 2a). Optimal oil degradation was achieved at broad N/P ratio between 5:1 and 20:1 with maximum value at 15:1 N/P ratios less than 1:1 or more than 20:1 resulted in retarded oil consumption.
Effect of iron: Culture medium with N/P molar ratio of 15:1 was supplemented with different concentrations of iron to investigate its role in biodegradation (Fig. 2b). Fe-free medium resulted in dramatic decrease in oil consumption, while addition of Fe stimulated the biodegradation process with maximum oil consumption observed in medium containing around 2.5 μ MFe.
Effect of salinity: Oil degrading cultures were supplemented with different salts in varying concentrations to estimate their salinity tolerance and oil consumption capacity. Addition of salt stimulated the biodegradation activity of the marine oil-enriched culture. At total salt concentrations between 3 percent and 7 percent enhanced oil consumption was observed with an optimum at 5 percent total salt. A dramatic decrease in oil consumption reaching 75 and 50 percent 15 and 10 percent total salt concentration, respectively, was observed if compared with the culture containing 5 percent salt (Fig. 2c).
Effect of Tween 20 concentrations: Aerated cultures containing 3 percent total salt and different concentration of Tween 20 (Fig. 2d) were incubated at optimal conditions. Culture receiving 0.05 g percent Tween showed about seven fold increase in oil consumption over control cultures lacking Tween. Increasing tween concentration over 0.1 percent retarded oil consumption. Aerated cultures with no Tween showed low oil consumption of about 8 percent.
Effect of temperature: The dependence of oil consumption by bacterial culture on incubation temperature (Table 4) was examined at standard incubation conditions. Maximum, oil consumption was reached at 35°C while negligible amounts of oil biodegradation were noticed at 5°C. Nearly the same amounts of oil were consumed at 25 and 45°C. Uninoculated flasks incubated at 45°C served as control for non biological oil loss by evaporation of the volatile fractions at this temperature.
Effect of inoculum density: Flasks containing 2.8 g percent oi1 were inoculated with inocula of increased optical density and incubated at standard conditions. It was clear, from the amount of recovered oils, that flasks receiving nearly five-fold the standard inoculum showed only about 15 percent increase in the amount of oil consumption (Table 4).
Compositional changes of the recovered oils: The compositional variations with respect to C-, H-, N-, S- and P - contents as well as C/H ratio of biodegraded recovered oils are given in (Table 5). Concerning the variation of C-content of the recovered marine oil in presence of different nitrogen sources, a general trend is difficult to establish. In contrast, a clear decrease in C-content of the recovered land oil was observed. Concerning the H-content, a slight decrease occurred in case of marine oil while land oil behaved differently showing an increase in H-content. The variation of the two oils was also clear with respect to P-content, where marine oil showed 1.5 to about 2 fold increase whereas land oil showed a general decreasing trend of P-content. Both oils behaved similarly with respect to N and S-contents. A general trend of increased N-and S-contents was maintained in most cases. With respect to C/H ratio, a clear difference between the two oils was observed. Marine oil showed increased C/H ratio, while land oil behaved oppositely showing decreased C/H ratio.
| Table 4: | Percentage of marine oil consumption by enriched bacterial culture incubated at standard conditions-affected by incubation temperature and inoculum density |
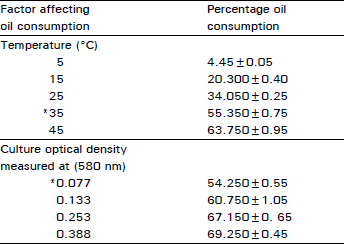 | |
DISCUSSION
Hydrocarbon degradation by microbial communities depends on the composition of the community and its adaptive response to the presence of hydrocarbons. Bacteria is a key agent of degradation which dominate in marine ecosystems. In the present work, trials to isolate natural bacterial flora from marine oil wells at Red Sea led to the isolation and identification of four genera, being Micrococcus sp., Bacillus sp., Corynebacterium sp. and Pseudomonas sp. Similar isolations from land oil sources were not successful. Hence, a mixed culture of such bacteria was used in the present biodegradation studies of both light crude oils. The results presented in this study show the efficiency of a natural bacterial enrichment culture in oil bioremediation in a closed system. In nature-open system-this process is greatly affected by the environmental factors which in turn affect the microbial activity to utilize spilled oils. The high sulfur contents in both oils could be attributed either to the origin of these oils from sediments of high sulfur content where anaerobic sulfate reduction predominates, or to previous biodegradation process of these oils in reservoir or during migration where non sulfur-bearing hydrocarbons are metabolized and hence oil tends to be more sulfurous (Waples, 1985). The ability of the enriched culture to utilize crude oils as sole carbon and energy source in presence of different nitrogen sources indicates its diversified metabolic capability. The preferential use of urea by this mixed culture Was accompanied by enhanced oil consumption, a feature which was reported for pure cultures of Pseudomonas ovalis and Candida tropicalis growing on petroleum hydrocarbons (Zaki and el-Badrawy, 1981 and Zaki et al., 1983).
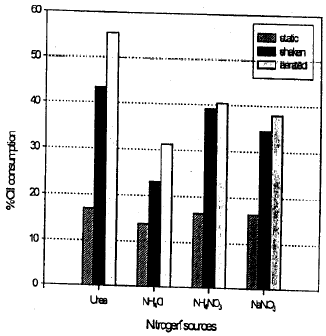 | |
| Fig. 1: | Marine oil consumption in static, shaked and aerated bacterial culture in presence of different nitrogen suruces |
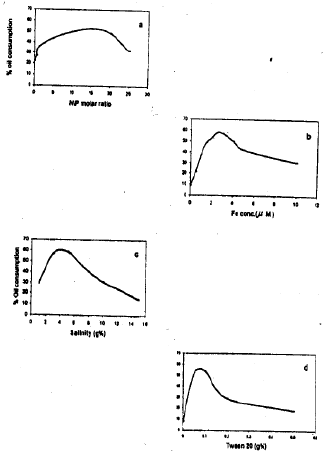 | |
| Fig. 2: | Effect of some environmental factors on oil consumption, a) Effect of NIP ratio, b) Effect of iron c) Effect of salinity d) Effect of Tween 20 concentration |
A pronounced decrease in oil biodegradation was observed when ammonium was associated with sulfate or phosphate. Atlas (1981) reported an inhibition of bacterial growth during oil biodegradation in presence of high phosphate concentrations. This may explain the lower values of oil consumption in the present study when N/P ratios are less than 5:1. One the other hand, Sen and Chatterjee (1983) and Sabek and Abu Zinada (1986) reported the preferential use of ammonium sulfate by Azotobacter and Methylomonas spp. growing on hydrocarbons.
The low amount of oil-consumed in absence of different external nitrogen sources and the detection of traces on NO2 in the medium, provided evidence for the ability fo the enriched bacterial culture to mineralize part of the organic oil nitrogen. However, participation of intracellular nitrogen of lysed bacterial cells as nitrogen source in these cultures may be an analternative explanation.
The release of hydrocarbons into aquatic environment which contain low concentrations of inorganic nutrients often produces high carbon/nitrogen or carbon/ phosphorus ratios, or both, which are unfavorable for microbial growth (Atlas, 1981 and Cooney, 1984). Adjustment of carbon/nitrogen/phosphorus ratios by the addition of nitrogen and phosphorus in the form of oleophilic fertilizers, including paraffinized urea, octylphosphate, magnesium ammonium phosphate and ferric octate, stimulates, the biodegradation of oils in open aquatic systems (Atlas and Bartha, 1973; Dibble and Bartha, 1976 and Olivieri et al., 1976). The carbon-content of the oils used in our study was about 85 percent (Table 1). Accordingly, at N/P ratio of 15:1 in presence of 2.8 g oil, maximum amounts of oils were consumed, i.e. C/N and C/P molar ratios of about 50:1 and 750:1 respectively, favored the bacterial growth and hence enhanced oil biodegradation. C/N and C/P ratios of about 101 and 100:1, 60:1 and 800:1, 20:1 and 100:1 were optimal ratios reported by Atlas and Bartha (1973), Dibble and Bartha (1979), Bridie and Bos (1971), respectively.
The metabolic availability fo petroleum iron and its contribution in the production of cell biomass resulted in low consumption of oil if compared with exogenously added iron. However, the utilization of intracellular iron in the course of population turnover furnishes an equally possible explanation for the low oil consumption. in iron-free medium. Studies by Dibble and Bartha (1976) on petroleum biodegradation in sea water, sowed that in less iron-rich sea water (1.2 μ M Fe) a pronounced decrease in oil consumption was observed if compared with the amount of consumed oil in iron-rich polluted sea water (5.2 μ M Fe). regardless of the nitrogen source present, aerated cultures were the most active oil consumers. This means that the catabolic oxidative power needed for degrading the vast number of hydrocarbons present in petroleum could be achieved with aeration, i.e. the maximal solubility of oxygen (7.1 mg 02/1 at 35°C under pressure of 1013 mbar) was maintained under the forced aeration condition employed in these experiments.
| Table 5: | Elemental analysis of recovered oil from aerated cultures containing different nitrogen sources |
 | |
In shaked cultures, the consumed oxygen was not replaced fast enough by diffusion from air because it was poorly soluble in water, hence, lower amounts of oils were consumed in these cultures.
A moderate salinity value between 3-7 percent total salt favored oil consumption. The water activity in cultures containing such amounts of salts is around 0.97 (Ward and Brock, 1978). It is known that microorganisms found in the sea usually have a specific requirement for the sodium ion (for cell membrane stability and activity of some enzymes), in addition to living optimally at the water activity of seawater which is a bout 0.98. The marine origin of the culture used in this study matches with these results (Brock and Madigan, 1991).
Microbial degradation of oils requires the presence of a large interfacial area of oil permitting better microbial contact to oils and faster in and out-diffusion of gases especially oxygen. This could be achieved by chemical agents as artificial dispersant or by biological agents as immobilized bacterial technology ad natural biosurfactant, or by physical agents as shaking or wave and wind action which break down oil slicks forming emulsion.
In static cultures lacking artificial dispersant, low amounts of oil were consumed. The natural production of biosurfactant by this culture may contribute in the utilization of oils in this case. Shake culture showed two-fold increase in the amount of consumed oils perhaps because of the cumulative effect of biosurfactant and shaking which resulted in better oil dispersion. The artificial dispersant of oil resulting in an enhanced oil consumption. However, increased concentrations of the added tween, more than 0.1 percent retarded oil biodegradation due to its toxicity to bacterial growth and metabolism. Increasing cell density of inoculum by five-fold resulted only in 15 percent increase in the amount of consumed oil. Nutrient deficiency is not the limiting factor. Production fotoxic by-or end-products or complete exhaustion of the metabolizable fraction of oil might be the reasons for this low oil consumption efficiency by heavily inoculated. cultures. Marine oil supplemented cultures containing optimum, nutrient concentrations and incubated at optimum environmental conditions consumed about 2/3 the amount of oil added in one week. This result may support the assumption of a limited amount of metabolizable fractions in the oil, i.e. about 30 percent of the oils are constituted of recalcitrant compounds.
Biodegradation changes the chemical and physical properties of oils. It is therefore important to understand the effect of biodegradation on oil composition. The two biodegraded oils showed increased N- and S-contents, a feature which is common for some biodegraded oils. This adds further support to the observations of Bailey et al. (1973) and Winters and Williams (1969). In heavily biodegraded oils, the sulfur content may increase. Waples (1985) and Wolcott et al. (1989) showed that the relative percentage of NSO- compounds increased with increasing biodegradation. The increase in both N- and S-contents of biodegraded oils was mainly attributed to the preferential microbial removal of hydrocarbons from oils leaving N- and S- contents of biodegraded oils was mainly attributed to the preferential microbial removal of hydrocarbons from oils leaving NSO-compounds preserved, hence their percentage increased in the recovered oils. In deep reservoirs, oil sulfur-content may increase as a result of sulfate reduction by sulfate-reducing bacteria.
Biodegraded marine oil samples showed elevated C/H ratio as a result of decreased H-content. This may reflect an increase in the relative degree of chain branching and increased carbon condensation with higher degree of unsaturation or aromaticity. Land oil samples behaved oppositly showing increased H-content, hence reduced C/H ratio which indicate higher degree of saturation. These results give an evidence for the variability fo the metabolizable fraction in both oils. A similar evidence has been reported for biodegraded oils by Wolcott et al. (1989). The presented data showed how the bacterial metabolic activities for crude oil biodegradation re affected by both the environmental and nutritional factors. Optimization fo these conditions resulted in enhanced oil biodegradation. Recovered weathered oil showed altered composition with respect to C-, H-, N-, S- and P-contents. Although a number of rate-limiting factors have been elucidated, the interactive nature of microorganisms, oil and environment still is not completely understood. Further study of oil biodegradation with special emphasis on -the problem of oil pollution is necessary for oil cleanup.
ACKNOWLEDGEMENT
Sincere appreciation is expressed to Prof. Dr. Abdel-Aziz Mahmoud Salama, Professor of Microbiology, in Botany Department, Faculty of Science, Cairo University, for critical reading of the manuscript and helpful discussions.
REFERENCES
- Atlas, R.M. and R. Bartha, 1972. Degradation and mineralization of petroleum in sea water: Limitation by nitrogen and phosphorous. Biotechnol. Bioeng., 14: 309-318.
CrossRefDirect Link - Atlas, R.M., 1981. Microbial degradation of petroleum hydrocarbons: An environmental perspective. Microbiol. Rev., 45: 180-209.
PubMedDirect Link - Atlas, R.M. and R. Bartha, 1973. Stimulated biodegradation of oil slicks using oleophilic fertilizers. Environ. Sci. Technol., 7: 538-541.
CrossRefDirect Link - Bailey, N.J.L., A.M. Jobson and M.A. Rogers, 1973. Bacterial degradation of crude oil: Comparison of field and experimental data. Chem. Geol., 11: 203-221.
CrossRefDirect Link - Barkay, T. and H. Pritchard, 1988. Adaptation of aquatic microbial communities to pollutant stress. Microbiol. Sci., 5: 165-169.
PubMedDirect Link - Dibble, J.T. and R. Bartha, 1979. Effect of environmental parameters on the biodegradation of oil sludge. Applied Environ. Microbiol., 37: 729-739.
Direct Link - Dibble, J.T. and R. Bartha, 1976. The effect of iron on the biodegradation of petroleum in seawater. Appl. Environ. Microbiol., 31: 544-550.
Direct Link - Leahy, J.G. and R.R. Colwell, 1990. Microbial degradation of hydrocarbons in the environment. Microbiol. Mol. Biol. Rev., 54: 305-315.
Direct Link - Milner, C.W.D., M.A. Rogers and C.R. Evans, 1977. Petroleum transformations in reservoirs. J. Geochem. Exp., 7: 101-153.
CrossRefDirect Link - Olivieri, R., P. Bacchin, A. Robertiello, N. Oddo, L. Degen and A. Tonolo, 1976. Microbial degradation of oil spills enhanced by a slow-release fertilizer. Applied Environ. Microbiol., 31: 629-634.
Direct Link - Rodriguez-Valera, F., F. Ruiz-Berraquero and A. Ramos-Cormenzana, 1980. Short communication Isolation of extremely Halophilic bacteria able to grow in defined inorganic media with single carbon sources. Microbiology, 119: 535-538.
CrossRefDirect Link - Sassen, R., 1980. Biodegradation of crude oil and mineral deposition in a shallow Gulf Coast salt dome. Org. Geochem., 2: 153-166.
Direct Link - Swannell, R.P., K. Lee and M. McDonagh, 1996. Field evaluations of marine oil spill bioremediation. Microbiol. Rev., 60: 342-365.
Direct Link - Walker, J.D., L. Petrakis and R.R. Colwell, 1976. Comparison of the biodegradability of crude and fuel oils. Can. J. Microbiol., 22: 598-602.
Direct Link