
Esmat E. A. Elwy Botany
Botany Department of Faculty of Science, Cairo University, Egypt
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 4 | Page No.: 1168-1172







ABSTRACT
Heavy metals significantly inhibited ascospores germination, differentiation and ascus formation in Dipodascopsis uninucleata. Sliver nitrate was the most inhibitory one even at low concentration (50 ppm). Among the tested zinc salts, zinc acetate was the most effective in inhibiting growth activities. Two phenomena have been observed in the present investigation. At concentration 600 ppm of manganese nitrate, D. uninucleata grow in the form of spherical pellicles with imperfect stage of reproduction represented by conidial heads similar to that of aspergilli. High concentration (1000 ppm), produced the yeast phase of D. uinucleata.
PDF Abstract XML References Citation
How to cite this article
Esmat E. A. Elwy Botany, 1999. Effect of Certain Heavy Metals on Dipodascopsis uninucleata. Pakistan Journal of Biological Sciences, 2: 1168-1172.
DOI: 10.3923/pjbs.1999.1168.1172
URL: https://scialert.net/abstract/?doi=pjbs.1999.1168.1172
DOI: 10.3923/pjbs.1999.1168.1172
URL: https://scialert.net/abstract/?doi=pjbs.1999.1168.1172
INTRODUCTION
Dipodascopsis uninucleata has a simple mycelial system. Biggs (1937), described the life cycle which starts by the germination of ascospores to form short mycelia, no asexual reproduction while sexual reproduction takes place by fusion of isogamous gametangia formed on 2 adjacent cells given rise to multispored asci.
On studying the effect of nutrients, radiation and different inhibitors on morphogenesis of D. uninucleata, it was found that vegetative growth and sexual reproduction were strictly related and any reduction or inhibition of ascus formation was always associated with similar effect on vegetative growth, except with N-methyl-N-nitro-N-nitrosoguanidine (NTG) which inhibited sexual reproduction and produced sterile mutant have the same pattern of growth as the wide type with larger cells and nuclei (Elwy, 1981).
Heavy metals may suppress or stimulate fungal responses according to their type and concentration (Ross, 1982). Lokesha and Somashekar (1990), reported some effects of heavy metals on mycelial growth of some fungi. Niwas et al. (1987) found that lead decreased significantly phylloplane fungi of wheat but did not affect Aureobasidium pullulans.
Some of the heavy metals may participate in the morphogenesis of a wide range of microorganisms. Zinc metal was reported to have a role in the morphogenesis of some fungal species e.g. Histoplasma capsulatum (Pine and Peacock, 1958), Mucor rouxii (Bartnicki-Garcia and Nickerson, 1962) and Candida albicans (Widra, 1964). Also, nitrate was found to favour mycelium to yeast transition more than ammounium in Aureobasidium pullulans (Park, 1982). The present investigation was carried out to study morphogenesis in the fungus Dipodascopsis uninucleata in response to various zinc salts as well as nitrates fo lead, manganese and silver.
MATERIALS AND METHODS
The strain of Dipodascopsis uninucleata Batra and Millner (Bigss) employed in this study was obtained from C.A.B. International Mycological Institute (IMI 86676) and maintained on MYGP medium (5 g glucose, 5 g peptone, 3 g yeast extract and 3 g malt extract in one liter distilled water) at 25 ± 1°C and 150 rpm in an orbital shaker.
Effect of zinc salts on growth and reproduction: Sets of triplicate flasks containing 50 ml MYGP were prepared and supplemented with the following zinc salts: chloride, sulphate, acetate, carbonate and phosphate as well as zinc metal. The following concentrations were used 10, 25, 50, 100, 200, 400 and 600 ppm. Control set was kept free of zinc or its salts. Culture flasks were inoculated with ascospores, to give a final concentration of 1x106 spore ml–1. Flasks were incubated at 25±1°C in an orbital shaker at 150 rpm. Samples were taken after 20 h incubation to examine ascospores germination; growth and reproduction were investigated in 48 h incubation using light microscope.
Effect of different metal nitrates on growth and reproduction: In this experiment manganese, lead and silver nitrates were used in the following concentrations: 50, 100, 200,400, 600 and 1000 ppm. The same procedures of inoculation and incubation were carried out as in the previous experiment. The medium used was MYG with the nitrate as the nitrogen source instead of peptone. Control sets were used using complete MYGP medium germination, growth, reproduction as well as morphological changes were observed and examined in 48 h incubation. Experiments were carried out in duplicate and the data presented as the mean value.
Statistical analysis: All data are given as the Mean±standard deviation and the difference between the means was tested using the "T" test at 95 percent confidence limit.
RESULTS
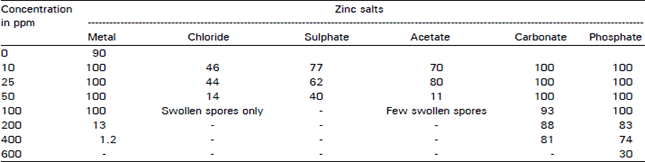
Effect of zinc metal and its salts on ascospores germination: Table 1 shows the effect of various concentrations fo zinc and its salts on spore germination of D. uninucleata. The results showed clearly that acetate, chloride and sulphate salts of zinc were the most effective and inhibited germination completely when used at concentration level of 100 ppm and higher.
| Table 1: | Effect of zinc metal and various zinc salts on germination percentage of D. ninucleata after 20 h |
 | |
| Table 2: | Effect of zinc metal and various zinc salts at different concentrations on the number of cells/hypha (A) and the percentage of successful trials (B) in D. uninucleata |
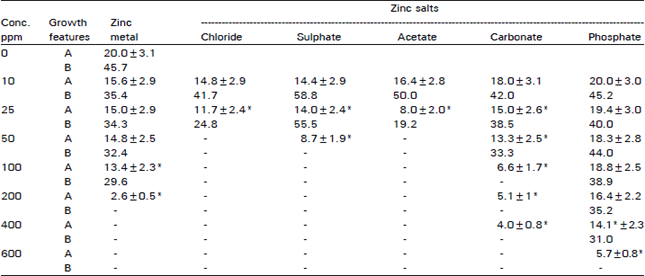 | |
| ±S.D: *Significant difference with the control at 95% C.L | |
| Table 3: | The effect of various concentration of manganese, lead and silver nitrates on the number of cells/hypha (A) and the percentage of successful trials (B) in D. uninucleata |
 | |
| ±S.D: *Significant difference with the control at 95% C.L | |
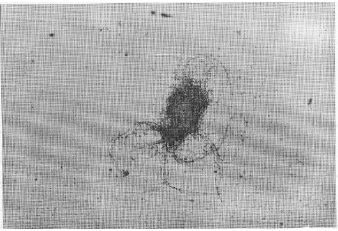 | |
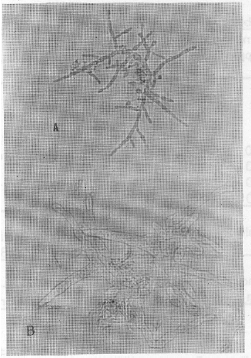
| Fig. 1: | Culture of D. uninucleata grown in the presence of 600 ppm manganese nitrate. It showed pellicle formation |
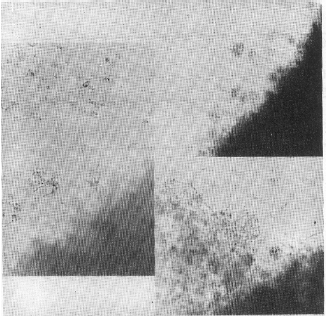 | |
| Fig. 2: | Magnified pellicle showing normal asci and hyphae of D. uninucleata intermingled with conidial head |
 | |
| Fig. 3a: | Culture of D. uninucleata grown with 1000 ppm managanese nitrate, showing branched myclelium with blasto spores. |
b: | Culure of D. uninulceata grown without manganese nitrate, showing branched mycelium with asci, ascospores and fused gametangia |
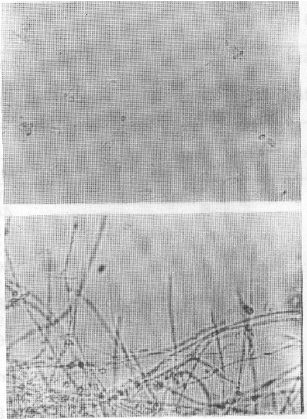 | |
| Fig. 4: | Pseudomycelium and yeast cells of D. uninucleata |
Zinc metal followed the previous mentioned salts but its inhibitory effect started at 200 ppm (13% germination) and 400 ppm inhibited the germination process. Zinc carbonate and zinc phosphate induced germination at low concentrations (100 ppm), then significant decrease in germination percentage was observed by the increase in concentration and complete inhibition has been achieved above 400 ppm.
Effect of zinc metal and its salts on growth and reproduction: Table 2 shows clearly the inhibitory effect of acetate, chloride and sulphate salts of zinc and this proved the observations in the previous experiment. Such inhibitory effect may be due to a direct effect on DNA or indirectly by inhibiting the activity of enzymes involved in growth and differentiation. Considering growth, low concentration (10 ppm) decreased the number of cells/hypha to 14-16 cellslhypha compared to 20 cells/hypha for the control. Concentration of 100 ppm and higher completely inhibited growth. Zinc metal was also effective and very short hyphae (two cells/hypha) was observed at 200 ppm, while that of the control was 20 cells/hypha. Complete inhibitory effect has been observed at 400 ppm.
Zinc phosphate was not effective as other salts. It produces hyphae of 14 cells at concentration level 400 ppm then decreased to 5.7 cells at 600 ppm compared to 20 cells/hypha for the control cultures. Incomplete inhibitory effect by phosphate salt, may be due to its utilization by fungal cells in many biochemical reactions involved in glucose phosphorylation and ATP synthesis. Zinc carbonate also produced significant effect at concentrations higher than 50 ppm and completely inhibited growth at 600 ppm.
Table 2 shows also the effect of zinc metal and its salts on sexual reproduction represented by the percent of successful trials of gametangial fusion. Acetate, chloride and sulphate salts of zinc were the most effective and concentrations higher than 25 ppm completely inhibited sexual reproduction. However, low concentrations of zinc sulphate (10, 25 ppm) induced gametangial fusion. Phosphate salt was also inhibitory and 400 ppm were required to reduce successful trials to 31 percent compared to 45.7 percent- the control. Zinc carbonate inhibited completely ascus formation at concentrations higher than 50 ppm.
Effect of nitrates of lead, manganese and silver on growth and reproduction: Silver nitrate significantly inhibited germination and 25 percent treated ascospores were able to germinate and grow to form short hyphae of 4-5 cells compared to 20 cells/hypha of the control (Table 3). In the developed hyphae, there was unsuccessful trials for gametangial fusion at low concentration (50 ppm).
Nitrates of manganese and lead had lower toxic effect than that of silver nitrate. This indicates that the metal part is the most important factor in determining the effect of salts on fungal activities. High concentration of manganese and lead (400 ppm) produced short hyphae of 6 and 9 cells respectively, while that of the control was 20 cells/hypha. Increasing the concentrations above 200 ppm for manganese and 400 ppm for lead significantly reduced successful trials.
The most important observation in the present study was in the case of manganese nitrate. At concentration 600 ppm, pellicle forme of growth appeared in the cultures (Fig. 1). Microscopic examination of the pellicles showed non-septated hyphae with conidial heads similar to that of aspergilli and were intermingled with the true growth of D. uninucleata (Fig. 2). Re-inoculation of pellicles into fresh MYGP medium produced pure cultures of the newly formed hyphae with conidial heads. At concentration 1000 ppm, pseudomycelia were observed in 48 h of growth. The survived ascospores developed in to short-branched hyphae on which blastospores were arranged randomly at right angles from the vegetative hyphae (Fig. 3) of pseudomycelia and yeast cells (Fig. 4).
DISCUSSION
Heavy metals have great effect on fungal activities and differentiation. The present investigation discussed their effect on germination, growth, sexual reproduction and morphogenesis of D. uninucleata. The inhibitory effect of the tested heavy metals salts has been observed clearly especially when applied in high concentrations. However, some metals such as silver showed its inhibitory effect at very low concentration (50 ppm), which is similar to that of Slade and Pegg (1993), who reported that Ag+ was the most toxic to zoospores of Phytopthora Nicotiana parasitica. Wainwright and Grayston (1983) also recorded toxicity of silver and lead nitrates. On the other hand, Variable effects of zinc and lead were also reported by Pachlewski and Chrusciak.(1986).
Slight induction has been observed with low concentrations of some metal salts as shown in the results. Similar results obtained by Dubey and Dwivedi (1988), who found that Zn at concentration 100 m μg/ml enhanced mycelial dry weight of Macrophomina phaseolina, while high concentration (500 m μg/ml) inhibited growth significantly. In addition, El-Sharouny et al. (1988), reported that counts of soil fungal population increased in some cases and reduced in others when treated with zinc or copper sulphates. The results of Lokesha and Somashekar (1990) were different from the present results, they reported that high concentrations of Zn supported growth of Drechslera halodes, D. tetramera and Curvularia lunata.
It is clear that the anion part of zinc salt has a role in its effect on growth and reproduction in D. uninucleata. The inhibitory effect of the used salts are in the following arrangement: acetate > chloride > sulphate > carbonate > phosphate.
Lead nitrate significantly reduced fungal population. Similarly, McCreight and Schroeder (1982) found that 200 ppm of lead arrested 5 spp. out of nine ectomycorrhizal fungi. It is evident that when vegetative growth was significantly affected a similar effect on sexual reproduction was observed, this is in agreement with Elwy (1981) regarding the effect of other substances on morphogenesis of D. uninucleata.
However a very interesting observation was recorded when MnNO3 was used at (600 and 1000 ppm). The formation of a morphological mutant which formed pellets and multiply asexually by condition and which is completely different form wild type, this support the observations of Byrne and Ward (1987), who found that the addition of divalent cations induced pelleting in some fungi. Such pellicle formation may be achieved by overcoming electrostatic repulsion between spores and mycelia.
This could be explained by the fact that the morphology of fungal cultures may be controlled by growth medium and environmental conditions (Calam, 1976). Therefore, when mycelia was packed in pellet structure, the decrease in aeration and diffusion of nutrients stimulated this form of growth and reproduction.
Several types of morphological mutant with abnormal sporulating structures were found in Aspergilli and Neurospora which may lose the capacity to sporulate normally even in favorable environmental conditions (Ross, 1982).
At higher concentration of manganese nitrate there was a transition from perfect stage of the wild type to yeast phase. This starts by elongation of ascospores to form long mycelia with blastospores, the growth of the yeast form on MYG, MYGP and MYG+MnNO3 produced a mixture of unicellular cells and long thin branched mycelia.
As morphogenesis and alterations in cell shape require integration of many cellular functions which occur in response to environmental signals MnNO3 induced such changes in the metabolic pathways of the wild type and produced the morphological mutant which was blocked at a certain stage of its sexual morphogenesis.
In conclusion, the present observation prove the hypothesis of Van Der Walt (1970) who proposed that Kluyveromyces polysporus might have been derived from Dipodascopsis uninuleata by regression of mycelium formation and gametangial sexuality on one hand and by the gain of blastospores formation on the other land.
REFERENCES
- Bartnicki-Garcia, S. and W.J. Nickerson, 1962. Nutrition, growth and morphogenesis of Mucor rouxii. J. Bacteriol., 84: 841-858.
Direct Link - Dubey, R.C. and R.S. Dwivedi, 1988. Effect of heavy metals on growth and survival of Macrophomina phaseolina (Tassi) Goid. Biol. Fertil. Soils, 6: 311-314.
CrossRefDirect Link - El-Sharouny, H.M.M., M.M. Bagy and A.A. El-Shanawany, 1988. Toxicity of heavy metals to Egyptian soil fungi. Int. Biodeterior., 24: 49-64.
CrossRefDirect Link - Byrne, G.S. and O.P. Ward, 1987. Effect of polymers on pelleting of Rhizopus arrhizus. Trans. Br. Mycol. Soc., 89: 367-371.
CrossRefDirect Link - McCreight, J.D. and D.B. Schroeder, 1982. Inhibition of growth of nine ectomycorrhizal fungi by cadmium, lead and nickel in vitro. Environ. Exp. Bot., 22: 1-7.
CrossRefDirect Link - Pachlewski, R. and E. Chrusciak, 1986. Effect of lead and zinc on the growth of some mycorrhizal fungi in vitro. Acta Mycol., 22: 73-80.
Direct Link - Park, D., 1982. Inorganic nitrogen nutrition and yeast-mycelial dimorphism in Aureobasidium pullulans. Trans. Br. Mycol. Soc., 78: 385-388.
CrossRefDirect Link - Ross, I.S., 1982. Effect of copper, cadmium and zinc on germination and mycelial growth in Candida albicans. Trans. Br. Mycol. Soc., 78: 543-545.
CrossRefDirect Link - Slade, S.J. and G.F. Pegg, 1993. The effect of silver and other metal ions on the in vitro growth of root‐rotting Phytophthora and other fungal species. Ann. Applied Biol., 122: 233-251.
CrossRefDirect Link - Wainwright, M. and S.J. Grayston, 1983. Reduction in heavy metal toxicity towards fungi by addition to media of sodium thiosulphate and sodium tetrathionate. Trans. Br. Mycol. Soc., 81: 541-546.
CrossRefDirect Link - Widra, A., 1964. Phosphate directed YM variation in Candida albicans. Mycopathol. Mycol. Applic., 23: 197-202.
CrossRefDirect Link