
Ali H. Bahkali
Not Available
Department of Botany and Microbiology, College of Science, King Saud University P.O. Box 2455, Riyadh 11451, Saudi Arabia
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 4 | Page No.: 1130-1134







ABSTRACT
The growth of phytopathogenic fungus Verticillium tricorpus on 15 carbohydrate substrates was monitored to analyse the range of polysaccharide-degrading enzymes and glucosidases production. The secretions of endo and exoenzymes, capable of degrading cellulosic, hemicellulosic and pectinolytic polysaccharide were observed during the course of the experiment. Pectinolytic activities were produced constitutively on all of the substrate tested, while cellulolytic enzymes were to induced in simple sugar (i.e., glucose or xylose) media. Polysaccharide growth substrates and cellulase inducers increased all of the enzyme activities tested.
PDF Abstract XML References Citation
How to cite this article
Ali H. Bahkali, 1999. The Production of Polysaccharide Degrading Enzymes By Phytopathogenic Fungus Verticillium tricorpus. Pakistan Journal of Biological Sciences, 2: 1130-1134.
DOI: 10.3923/pjbs.1999.1130.1134
URL: https://scialert.net/abstract/?doi=pjbs.1999.1130.1134
DOI: 10.3923/pjbs.1999.1130.1134
URL: https://scialert.net/abstract/?doi=pjbs.1999.1130.1134
INTRODUCTION
The ability of many organism to cause disease and in some cases death, of certain plant may be of great economic consequence. Verticillium species are known to attack a wide variety of economically important crops. Of these V. albo-atrum Reink and Berth., V. dahliae Klebahn and V. tricorpus Isaac are of great value and most of the research has been concentrated on these species (Isaac, 1967; Bahkali, 1983, 1987).
Most phytopathogenic fungi are capable of producing polysaccharide-degrading enzymes which can alter or degrade a number of the polymeric carbohydrates found in higher plant cell walls (Cooper and Wood, 1980; Barash et al., 1984; Bahkali, 1983, 1987, 1989, 1992; Bahkali et al., 1997; Aljohimi, 1995). Some pathogens produced both pectic hydrolases and lyases (Goodenought and Maw, 1974; Cooper and Wood, 1980; Bahkali, 1987, 1992), while others produce one of these enzymes (Bateman, 1972). The ability of pathogens to produce multiple forms of polysaccharide-degrading enzymes also appears to be common. This multiplicity confers flexibility, increasing the efficiency of the hydrolytic complex. It is also suggested that the existence of these enzymes would enhance the probability that a given substrate would be more effectively degraded (Cooper and Wood, 1980).
Extracellular proteins secreted by V. tricorpus are able to macerate tissues and degrade cell wall components. They must thus contain all of the enzymes corresponding to the types of glycosidic linkages present in the cell wall polysaccharide. Verticillium tricorpus is known to produce pectinolytic and cellulolytic enzymes (Bahkali, 1987, 1995). The level of these enzymes the diversity of polysaccharide produced by V. tricorpus and the mechanisms controlling expression of cell wall-degrading enzymes are poorly understood.
The objective of this study was to examine in detail the range of polysaccharide depolymerases and glucoside hydrolases secreted by V. tricorpus. Cultures were grown on a range of monosaccharides, disaccharides and polysaccharide so that the effects of the available carbon sources on enzyme production and activity could evaluated.
MATERIALS AND METHODS
Chemicals: The polysaccharide used as the carbon source and as enzyme substrates were cellulose microcrystalline (Avicel) from Merck (Darmstadt, Germany), hydroxymethyl cellulose (HEC), apple pectin (esterification, 70 to 75%) from Fulka (Buchs, Switzerland), Laminarin (Larch Wood) and arabinogalactan from Sigma (St. Louis, Mo.) and xylan (oat splets), Na+ polygalacturonic acid (Mr, 25,000), carboxymethyle cellulose (CMC; degree of polymerization, 500 to 520) and galactan from Serva (Heidelberg, Germany). All other reagents, glycosides used as carbon sources and nitrophenyl and methylumbelliferyl used as substrates were purchased from Sigma.
Culture conditions: V. tricorpus (isolate 227.84), obtained from Centraalbureau Voor Shimmel-cultures (Netherlands), was used throughout this investigation. The isolate was originally obtained from a wilted potato plant (Solanum tuberosum L.) and was maintained on slants of potato dextrose agar, stored at 4°C.
Verticillium tricorpus was grown on a liquid minimal medium supplemented with a 0.5 percent (wt/vol) carbon source. the minimal medium contained NH4NO3 (2 g/liter), KH2PO4 (1 g/liter), MgSO4 (0.1 g/liter), NaOH (1 g/liter) and DI-malic acid (3 g/litter). Cultures were maintained on potato dextrose agar. For enzyme production, 200 ml cultures inoculated with 20 plugs (4 mm diameter) removed from the growing edge of 4-day old colonies were grown for 7 days at 24°C under constant agitation. Cultures were harvested by filtration through Whatman No. 1 filter paper. Filtrates were dialyzed against distilled water overnight at 4°C and then freeze-dried. Cultures were harvested onto pre-weighted glass sheets and dried in an air oven at 70°C for 24 h. The dry weight of the mycelium mat was then calculated and fungal growth was expressed as mg dry weight per culture.
Enzymes methods: Freeze-dried culture media were solubilized in 4 ml 0.1 M Na+ acetate buffer, pH6 and then precipitated to 25 percent saturation of ammonium sulphate and centrifuged at 10,000 x g for 10 min. The supernatant was brought to 85 percent saturation of (NH4)2SO4 and then centrifuged.
Pellets dissolved in Na+ acetate buffer and dialyzed against distilled water and then against the Na+ acetate buffer were used as enzyme sources. Protein content was determined by the method described by Bradford (1976) with bovine serum albumin as the standard.
Glycoside hydrolase activities were determined by measuring the rate of P-nitrophenol released from the appropriate P-nitrophenyl derivatives (Table 1-4). The standard reaction mixture (1 ml) contained 20 μl of enzymes solution and 2mg of substrate dissolved in 0.1 M Na+ acetate buffer, ph6. After 15 min of incubation at 50°C, reactions were stopped by the addition of 3 ml of dinitro salicylic reagent (Miller, 1959). Tubes were placed in a boiling-water bath for 8 min. The A528 was red with appropriate single sugars as standards (Hebraud and Fevre, 1998).
Enzyme and substrate controls were included in all assays. All enzyme reactions were linear over the period of assay. Enzyme activities are expressed as nanomoles of P. nitrophenol or the equivalent micrograms of reducing sugar.
Pathogenicity test: Pathogenicity experiments in the green house were conducted on the "Ajax' cultivar of potato plant (Solanum tuberosum L.). This cultivar was chosen because of its susceptibility to Verticillium spp. found in the Kingdom of Saudi Arabia, where Verticillium wilt occurs in potato plants. It was likely, therefore that the pathogenic potential of V. tricorpus in potato would be determined best in this cultivar of in cultivars of similar susceptibility. The fungus was maintained on PDA slants as a wild type culture (produces all of the major morphological traits characteristic of the species in culture isolated from naturally infected plants). Conidial suspensions were made from 6-day-old cultures of the fungus with sterile distilled water and 0.5 ml, of the suspension were distributed over the surface of larger PDA slants to produce uniform lawn of growth. After 5 to 6 days of incubation at 25°C, another heavy conidial suspension were prepared from these slants in 10 ml. of sterile distilled water and diluted to produce approximately 6 x 106 viable canidia/nnl. Approximately 9 ml, of conidial suspension was inoculated to the root ball of five 6-weekold potato plants growing in a greenhouse-mix soil in 4 inch plastic pots. The control were five plants treated with sterile distilled water without fungus. All plants were held at 23°C. in a greenhouse. Observations for symptoms were begun 3 weeks after inoculation.
RESULTS AND DISCUSSION
Verticillium tricorpus induced wilt symptoms to potato plants. All the leaflets along one side of each rachis became chlorotic before complete necrosis took place. Eventually the remainder of the leaf became chlorotic and then died (Fig. 1). Isaac (1967) concluded that V. tricorpus was pathogenic to a narrow range of host plants and that external symptoms were apparent only after an extended period and symptoms were similar in the other host plants. Enzymes capable of degrading a wide range of glucosidase and polysaccharide were detected in cell-free culture supernatant. Growth of V. tricorpus on various polysaccharide used as the sole carbon sour, demonstrated that the fungus secretes enzymes that convert cellulosic, pectinolytic and hemicellulolytic substrates to assimilable simple sugars. The biomasses produced in these cultures were much lower than those obtained on glucose r xylose media (Table 1 and 2).
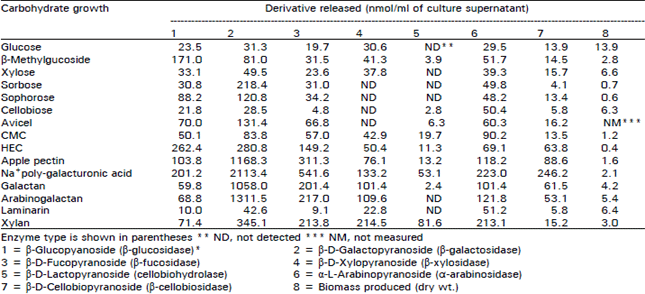
Glucose and xylose grow cultures exhibited nearly all of the enzyme activities tested except for xylanase and cellobiohydrolase, which were not detected. The levels of the other enzymatic activities were low except those for pectinase, polygalacturonase and β-1,3-glucanase, indication that glucose and xylose did not completely prevent synthesis of all of the enzyme activities and that some enzymes are formed constitutively.
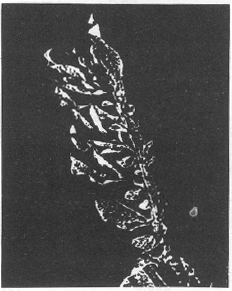 | |
| Fig. 1: | Typical unilateral wilting in the leaf of potato plant by Verticillium |
The abilities of various substrates to modify enzyme production were studied by growing V. tricorpus on polysaaccharides and sugars which are known to regulate enzymes tested were produced. In comparison with glucose medium, polygalacturonic acid medium showed the highest production of exoenzymes. Maximal pectinolytic activities were also measured in these media. Pectinase activities against apple pectin and polygalactruronic acid were higher in these media than those activities in glucose medium. Xylanase and β-1,4 glucanase activities which were not detected in glucose medium were recovered from these media. Different levels of exoenzymes activities, i.e., β-1,3 glucancase and pectinolytic enzymes, were present in media containing the cellulosic substrate crystalline cellulose (Avicel) and their soluble derivatives (HEC and CMC), but xylanase and endo β-1,4 glucanse activities were present only in HEC and CMC media. The secretion of extracellular proteins in the presence of pectinolytic substrates was a increased (Table 3). Among the hemicellulolytic polysaccharide used, Laminarin was very poor substrate for enzymes productions. In contrast, in medium containing xylan, the other hemicellulolytic substrate, all of the enzyme activities were present.
| Table 1: | Glycosidase activities of extracellular preparations from V. tricorpus grown on a range of carbon sources and expressed as amounts of P-nitrophenol released per milliner of culture supernatant |
 | |
| Table 2: | Polysaccharidase activities of extracellular preparation from V. tricorpus grown on a range of carbon sources and expressed as amounts of reducing sugar released per milliner of culture |
 | |
| * Enzyme type is shown in parentheses** ND, not detected*** NM, not measured | |
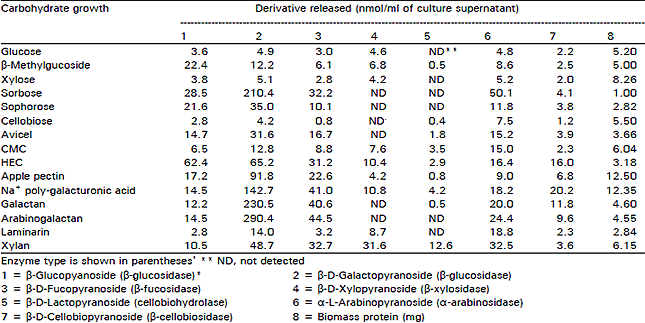
The induction of cellulolytic enzymes by crystalline cellulose or soluble derivative molecules which cannot enter the cell is held to be mediated by low molecular weight cellulose degradation products or their transglycosylation products (Messner et al., 1988). Several potent inducers in other organisms, i.e., sophorose L-sorbose (Kawamori et al., 1986), cellobiose (Khiyami, 1994) and β-methyl glucoside were used as the sole carbon source. Among these known cellulolytic enzyme inducers, only β-methyl glucoside and cellobiose were efficient for the production of β-1,4 glucanase activities degrading CMC and β-D-lactopyranoside and, surprisingly, of pectinase, polygalacturonases and β-1,3 glucanases. In contrast, sophorose and sorbose were poor substrates because notable increases occurred and in only a pectinase, β-glucosidase and β-galactosidase in sophorose grown cultures and in β-galactosidase in sorbose grown cultures as compared with the increases in glucose grown cultures. This lack of enzyme production by sophorse was noted by extracellular proteins since only a few amount of proteins were detected Table 3. When comparisons of enzyme production were made at the level of the specific activity (i.e., enzymatic activity per microgram of protein), maximal activities were generally obtained in culture grown on the appropriate structure related polysaccharide (Table 3 and 4). The highest specific activities of β-glucosidase, β-1,4 glucanase, β-galactosidase, β-1,3 glucanase and β- xylosidase were obtained, respectively, in HEC, arabinogalactan, laminarin and xylan grown cultures. However, increases of enzyme activity unrelated to the carbon source used were also frequently observed. Arabinosidase, cellobiosidase, cellobiohydrolase and pectinase had the highest specific activities in sorbose, plygalacturonic, acid and xylan grown cultures.
| Table 3: | Glycosidase activities of extracellular preparations from V. tricorpus grown on a range of carbon sources and expressed as nmole of P-nitrophenol released |
 | |
| Table 4: | Polysaccharidase activities of extracellular preparation from V. tricorpus grown on expressed as μg of reducing sugar released |
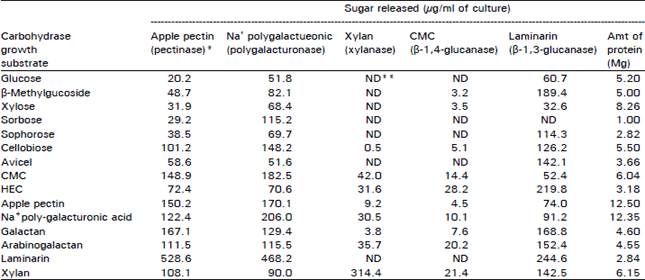 | |
| *Enzyme type is shown in parentheses ** ND, not detect | |
Previous studies on V. tricorpus have demonstrated its potential to degrade the principal plant structure polysaccharide (Bahkali, 1983, 1989, 1995). The data presented here confirmed that this pathogenic fungus has a very wide range of polysachride and glycosidase activities. This fungus possesses the glycoside hydrolase activities that complement the Polysaccharidase enzymes which are also formed, conferring an enzymatic potential to release monosaccharides from each plant cell wall polymer. Glycosidases may also remove side groups of heteropolysaccharides, facilitating the action of endoenzmes. A specificity of glycoside hydrolases has also been observed in other fungi; β-xylosidases from Trichoderma reesei and from Neurospora crassa exhibit α-arabinosidase and β-glucosidase activities, respectively.
The mechanism controlling expression of cell wall-degrading enzymes in fungi is not well understood. In culture, the production of hydrolytic enzymes by many fungi requires substrate and is repressed by preferred carbon sources such as glucose (Bahkali, 1983; Collmer and Keen, 1986; Keen et al., 1987).
The induction and repression of the hydrolytic enzymes were studied by growing V. tricorpus on a variety of carbon sources. The enzymes activities, except
those of pectinase and β-1,3 glucanase were low or absent in glucose grown cultures. The activities increased considerably when various polysaccharide were used as the carbon source. Pectinoiytic enzymes seem to be produced constitutively, while β-1,4 glucanases are induced by polysaccharide. Cellulolytic or pectinolytic substrates were able to induce cellulolytic and pectinolytic enzymes, indicating an aspecificity of the induction or a common regulatory system. This was also illustrated by the high pectinolytic activities of cultures obtained on cellobiose or methylgucoside, inducers of cellulolytic enzymes.
In the present work V. tricorpus produces polysaccharide depolymerases and glucosidases necessary to degrade the important structural cell wall polysaccharide, i.e., pectin, cellulose and hemicellulose. The secretion of this wide range of enzyme provides this pathogenic fungus with the ability to attack hosts which differ in their polysaccharide cell wall composition and could explain the lack of host specificity of this fungus. Because of the variety of enzyme produced, this fungus may also offer commercial potential. Optimization studies will serve to increase the understanding of factors that control the production, activity and consequently the role of the enzymes.
ACKNOWLEDGEMENT
The author gratefully acknowledges the support of the Research Center, College of Science. He also thanks Dr. M.A. Shinwari, of the Biochemistry Department of King Saud University for reading the manuscript.
REFERENCES
- Bahkali, A.H., 1992. In vitro production of pectolytic and cellulolytic enzymes by Colletotrichum lindemuthianum isolated from soybean grown in Saudi Arabia. World J. Microbiol. Biotechnol., 8: 55-59.
CrossRefDirect Link - Bahkali, A.H., 1995. Production of cellulase, xylanase and polygalacturonase by Verticillium tricorpus on different substrates. Bioresour. Technol., 51: 171-174.
CrossRefDirect Link - Bahkali, A.H., A.S. Al-Khaliel and K.A. Elkhider, 1997. In vitro and in vivo production of pectolytic enzymes by some phytopathogenic fungi isolated from Southwest Saudi Arabia. J. King Saud Univ., 9: 125-137.
Direct Link - Bateman, D.F., 1972. The polygalacturonase complex produced by Sclerotium rolfsii. Physiol. Plant Pathol., 2: 175-184.
CrossRef - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Cooper, R.M. and R.K.S. Wood, 1980. Cell wall degrading enzymes of vascular wilt fungi. III. Possible involvement of endo-pectin lyase in Verticillium wilt of tomato. Physiol. Plant Pathol., 16: 285-300.
CrossRefDirect Link - Collmer, A. and N.T. Keen, 1986. The role of pectic enzymes in plant pathogenesis. Annu. Rev. Phytopathol., 24: 383-409.
CrossRef - Hebraud, M. and M. Fevre, 1988. Characterization of glycoside and polysaccharide hydrolases secreted by the rumen anaerobic fungi Neocallimastix frontalis, Sphaeromonas communis and Piromonas communis. Microbiology, 134: 1123-1129.
CrossRefDirect Link - Kawamori, M., Y. Morikawa and S. Takasawa, 1986. Induction and production of cellulases by L-sorbose in Trichoderma reesei. Applied Microbiol. Biotechnol., 24: 449-453.
CrossRefDirect Link - Messner, R., F. Gruber and C.P. Kubicek, 1988. Differential regulation of synthesis of multiple forms of specific endoglucanases by Trichoderma reesei QM9414. J. Bacteriol., 170: 3689-3693.
CrossRef - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link