
Masood Akhtar
Department of Veterinary Parasitology, University of Agriculture, Faisalabad-38040, Pakistan
C.S. Hayat
Department of Veterinary Parasitology, University of Agriculture, Faisalabad-38040, Pakistan
Pakistan Journal of Biological Sciences
Year: 1998 | Volume: 1 | Issue: 4 | Page No.: 383-385







ABSTRACT
Optimum conditions for erythrocyte rosette formation (ERF) assay were determined to detect the cellular responses in immunized buffaloes against ticks. ERF with buffalo lymphocytes was maximum at 30°C for 30 minutes. The gentle handling of cell mixtures (lymphocytes and RBC) was recommended since binding forces between red cells and T-lymphocytes are week and easily disrupted. It was found that the buffalo lymphocytes did not bind to rabbit erythrocytes to form rosettes. The appearance of rosettes formed with sheep RBC was indistinguishable in appearance from rosettes formed with human erythrocytes. The percentage of E-rosettes formed with sheep RBC was slightly less (p>0.05) (mean 28.00±5.8, range 19- 38), than the percentage formed with human RBC (mean 31.90+7.7, range 17-45). The test was found to be quick, convenient and sensitive to detect the cellular responses in immunized buffaloes against ticks.
PDF Abstract XML References Citation
How to cite this article
Masood Akhtar and C.S. Hayat, 1998. Erythrocyte Rosette Formation Assay for the Detection of Cell Mediated Immune Responses in Buffaloes Vaccinated Against Ticks. Pakistan Journal of Biological Sciences, 1: 383-385.
DOI: 10.3923/pjbs.1998.383.385
URL: https://scialert.net/abstract/?doi=pjbs.1998.383.385
DOI: 10.3923/pjbs.1998.383.385
URL: https://scialert.net/abstract/?doi=pjbs.1998.383.385
INTRODUCTION
The cell mediated immune (CMI) response against bacterial and viral vaccines have been assayed by a number of methods including delayed hypersensitivity (Jones, 1974), macrophage and leukocyte migration inhibition (Silverman, 1970; Buening, 1973), lymphoblstogenesis (Swiderska et al., 1971), erythrocyte rosette formation (Mayr et al., 1982) and leukocyte adherence inhibition (Holan, 1982). However, only delayed hypersensitivity and lymphoblstogenesis have been employed to detect cellular response to antitick vaccines (Opdebeeck arid Daly, 1990). The earlier test is well characterized and classified method to detect cellular response in vaccinated animals. Therefore, certain in vitro assays have been widely used for detecting specific lymphocyte reactivity in animals without altering the immunological status. However, the injection of antigens into an animal may alter the immunological status and further evaluation of humoral or cellular responses may not be valid (Jones and Buening, 1983). Lymphoblstogenesis is a relatively more sensitive and successfully used technique to assay cellular response against ticks. The limitation of this method is long time (64-136 hours) to wait for results (Opdebeeck and Daly, 1990). The present study reports the standardization and application of erythrocyte rosette formation assay to detect cellular responses in vaccinated buffaloes against ticks.
MATERIALS AND METHODS
Experimental Animals: Ten each unvaccinated and vaccinated buffaloes with midgut, Boophilus microplus, cell culture vaccine (Akhtar, 1995) were used in the present study.
Isolation of lymphocytes: Buffalo lymphocytes were separated from the blood following the technique of Akhtar (1995). Briefly, 3 percent solution of gelatin (Sigma Laboratories, St. Louis, USA) in normal saline was sterilized by keeping in boiling water for 30 minutes daily for three consecutive days.
A 15 mL, gelatin solution containing 20 units/mL preservative free heparin was added to sterile syringes each containing an equal volume of anticoagulated blood collected from experimental animals.
Blood and gelatin were mixed properly and allowed to stand vertically at 37°C in an incubator for 30 minutes. Needles of the syringes were bent to an angle of 90° with a needle-guard. Plunger was pushed and supernatant containing lymphocytes was collected in sterile tubes and centrifuged (2000 rpm/15 minutes).
Lymphocytes pellet were washed twice and resuspended in 15 mL Minimum Essential Medium (MEM; Sigma Laboratories, St. Louis, USA) containing one per cent antibiotic (x100 antibiotics mixture of penicillin, streptomycin and amphotericin (B:GIBCO, Grand Island, NY, USA). Lymphocytes suspension was incubated at 37°C for 30 minutes to remove granulocytes.
Lymphocytes were counted by adding 0.1 mL sample from each aliquot in snap-cap tubes each containing 0.9 mL of one per cent trypan blue (Merck, Germany). Final count was adjusted to 5 x 106 lymphocytes per mL. Cell suspensions having at least 90 per cent viable lymphocytes were used for test.
Preparation of erythrocytes: A 10 mL blood each of human (group 0), sheep and rabbits were collected separately in heparinized syringe and centrifuged (1000 rpm/ 5 minutes). Plasma and huffy coat were discarded. Sedimented erythrocytes were washed twice in Phosphate Buffered Saline (PBS, pH 7.2) and stored at 4°C as a 50 per cent suspension in Alsevers solution. Erythrocytes were washed twice in PBS and resuspended and counted per mL in MEM before they were used for test.
Erythrocyte rosette formation assay: A 0.2 ml aliquot of cell suspension (1×106 lymphocytes) in MEM of unvaccinated animals was mixed with 0.2 mL of erythrocytes (12×106 cells) and incubated at room temperature (30°C) for 30 minutes. The cell mixtures were centrifuged (1000 rpm/ 5 minutes) and pellets were resuspended in MEM. A drop of each cell suspension was added to a hemocytometer and 200 lymphocytes were counted. The lymphocytes binding two or more erythrocytes were considered as rosette positive. Hence, per cent rosette formation was determined for each vaccine.
Under optimum conditions, erythrocyte rosette formation assay was performed with human erythrocytes and the blood lymphocytes of the experimental animals before and after vaccination. Per cent rosette enhancement was determined following the method of Bowles et al. (1975).
RESULTS
Optimum conditions for erythrocyte rosette formation were determined by using lymphocytes separated from unvaccinted buffaloes. Incubation of buffalo lymphocytes and human RBC mixture at 30°C for 30 minutes followed by centrifugation (100 rpm/5 minutes) resulted in a high percentage (37.2±6.6) of lymphocyte forming erythrocyte rosette. High percentages of rosettes were also formed after incubation of the lymphocytes RBC mixture at 37°C for 5 minutes followed by centrifugation (28.9±4.7) or 4°C for 5 minutes followed by centrifugation and further incubation at 4°C for 5 minutes 129.8±9.9) but the percentage of rosette lymphocytes at these times and temperatures was nonsignificant than when rosettes were formed at 30°C for 30 minutes (Table 1).
The percentage of lymphocytes which formed E-rosettes was not significantly different whether the lymphocytes were incubated in MEM or Hank’s balanced salt solution but there was a significant difference (p<0.05) in percentage of lymphocytes formed rosettes when incubated in PBS. The addition of Foetal calf serum (FCS) at 5 or 10 percent did not markedly alter the percentage of lymphocytes which formed E-rosette with human RBC.
It was found that the bovine lymphocytes did not bind to rabbit erythrocytes to form rosettes. The appearance of rosettes formed with sheep RBC was indistinguishable in appearance from rosettes with human erythrocytes. The percentage of E-rosettes formed with sheep RBC was slightly less, mean 28±5.8 (range 19 to 38), than the percentage formed with human RBC, mean 31.90±7.7 (range 17 to 45).
Erythrocyte rosette formation before and after 3 and 6 weeks vaccination was 37.00±1.90 and; 44.34±1.65 and 53.94±1.55 percent, respectively. The rosette enhancement 3 and 6 weeks post vaccination was 19.83 and 45.77 percent, respectively.
DISCUSSION
Erythrocyte rosette {E-rosette) formation assay has been successfully used to measure the CMI response produced due to different bacterial and viral vaccine. But so for the technique has not been evaluated to measure CMI response due to tick vaccines. In the present studies, peripheral blood lymphocytes from unvaccinated buffaloes were investigated to determine the optimum conditions for E-rosette formation with human RBC’s. ERF was significantly higher at 30°C for 30 minutes compared to that 37°C or 4°C for 5 minutes. The release of different soluble factors like lymphokines from T-lymphocytes during incubation acts as a part of binding force between erythrocytes and lymphocytes to form rosettes. Therefore, variation in temperature affects the release of soluble factors which may influence ERF.
The percentage of lymphocytes forming rosettes was significantly higher (p<0.05) when incubated in MEM or Hank’s balanced salt solution compared to that in PBS. This may be attributed to the presence of excessive amount of NaCl in PBS which has been reported to be an inhibitor of calcium ion bindings between the T-cells and erythrocytes required for rosette formation (Pang et al., 1976)
The addition of FCS at 5 or 10 per cent did not markedly alter the percentage of lymphocytes which formed E-rosettes with human RBC.
Handling of cell mixtures containing E-rosetted cells also found to have effect on per cent E-rosette enhancement. So the gentle handling of cell mixtures is recommended since binding forces between red cells and T-Iymphocytes are week and easily disrupted (Brain et al., 1970).
| Table 1: | Effect of temperature and time of incubation on rosette formation by buffalo lymphocytes and human RBC’s |
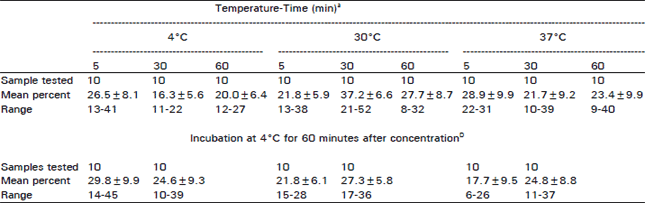 | |
| aBuffalo lymphocytes and human RBC’s were incubated at the specific time and temperature followed by centrifugation at 1000 spm for 5 minutes. Cell pellet was resuspended and rosette percentages were determined. DLymphocytes and RBC’s were incubated at 4,30 and 37°C for 5 or 30 minutes, centrifuged at 1000 rpm for 5 minutes and further incubated at 4°C for 60 minutes before resuspension of the cell pellet and rosette determination. | |
E-rosette formation was also attempted using erythrocytes of sheep and rabbits. It was found that the bovine lymphocytes did not bind to rabbit erythrocytes to form rosettes. The appearance of rosettes formed with sheep RBC was indistinguishable in appearance from rosettes formed with human erythrocytes. A striking feature of the bovine E-rosette was the comparative size of lymphocytes (mean) and human (mean 7.0u) or sheep (mean 4,4u) erythrocytes (Schermer, 1967), as contrasted to a mean 7.5u for rabbit erythrocytes, resulted into a limiting number of red cells making contact with lymphocytes. Because of the large red cells (rabbit) size and weak binding forces it may be possible that some of the rosettes were disrupted before the preparation was examined microscopically and were thus designated null cells.
A comparison was made between the percentages of lymphocytes from blood sample positive for erythrocyte rosette with human and sheep RBC’s. The percentage of E-rosettes formed with sheep RBC was slightly less (mean 28.00, (range 19-38), than the percentage formed with human RBC (mean 31.90±5.8, range 17-45).
Erythrocyte rosette formation assay was found to be quick, convenient and sensitive in vitro assay to detect the cellular responses interms of percent rosette enhancement in immunized buffaloes against ticks.
REFERENCES
- Bowles, C.A., G.S. White and D. Lucas, 1975. Rosette formation by canine peripheral blood lymphocytes. J. Immunol., 114: 399-402.
Direct Link - Brain, P., J. Gordon and W.A. Willetts, 1970. Rosette formation by peripheral lymphocytes. Clin. Exp. Immunol., 6: 681-688.
Direct Link - Buening, G.M., 1973. Cell-mediated immune responses in calves with anaplasmosis. Am. J. Vet. Res., 34: 757-763.
PubMed - Jones, R.L. and G.M. Buening, 1983. Brucella abortus antigen-induced lymphocyte reactivity in guinea pigs. Vet. Immunol. Immunopathol., 4: 433-443.
CrossRefDirect Link - Jones, L.M., 1974. Specificity of Brucella protein antigens and role of lipopolysaccharide antigens in eliciting delayed hypersensitivity reactions in sensitized guinea pigs. Annales Recherches Veterinaires, 5: 189-199.
Direct Link - Mayr, B., I. Vogel, W. Graninger, G. Schlerka, F. Wockl and W. Schleger, 1982. Circulating immune cells and immune complexes in peripheral blood of healthy and of bovine leukemia virus-infected cows and lymphosarcomatous calves. Vet. Immunol. Immunopathol., 3: 475-484.
CrossRefDirect Link - Opdebeeck, J.P. and K.E. Daly, 1990. Immune responses of infested and vaccinated Hereford cattle to antigens of the cattle tick, Boophilus microplus. Vet. Immunol. Immunopathol., 25: 99-108.
CrossRefDirect Link - Pang, G.T.M., P.M. Wiggins and J.D. Wilson, 1976. Nature of the union between sheep red blood cells and T lymphocytes. Int. Arch. Allergy Immunol., 51: 751-756.
CrossRefDirect Link - Silverman, M.S., 1970. The macrophage in cellular and humoral immunity. J. Reticuloendothelial Soc., 8: 105-123.
PubMed - Swiderska, H., T. Osuch and W.J. Brzosko, 1971. Peripheral blood lymphocyte blast transformation test as applied in the diagnosis of brucellosis. Medycyna Doswiadczalna Mikrobiologia, 23: 135-139.
PubMedDirect Link