
M. Aslam
Nuclear Institute for Agriculture and Biology Faisalabad-Pakistan
M. T. Elahi
Nuclear Institute for Agriculture and Biology Faisalabad-Pakistan
N. Iqbal
Nuclear Institute for Agriculture and Biology Faisalabad-Pakistan
Pakistan Journal of Biological Sciences
Year: 1998 | Volume: 1 | Issue: 4 | Page No.: 291-294







ABSTRACT
The research studies were carried out to enhance the incorporation of G. arboreum and G. barbadense genes for disease resistance and quality traits respectively into G. hirsutum through DNA-mediated embryo transformation technique. The self fertilized flower/ovaries of the recipient were injected with the donors DNA solutions irradiated at low doses ie. 2.5 Gy of gamma rays. The result of D1 and D2 generation revealed the enhanced incorporation and preservation of the donor parent traits into the recipient. Higher percentage of CLCV resistant plants and plants with increased quality traits were obtained from the irradiation DNA treatments. The transformed genotypes had higher yield and other economic traits better as compared to recipient. Moreover the expression of qualitative traits of the donor i.e. petal spot, pollen colour and flower colours were observed in D2 generation.
PDF Abstract XML References Citation
How to cite this article
M. Aslam, M. T. Elahi and N. Iqbal, 1998. Development of Improved Germplasm of Cotton Through Radiation and
DNA-mediated Embryo Transformation Technique. Pakistan Journal of Biological Sciences, 1: 291-294.
DOI: 10.3923/pjbs.1998.291.294
URL: https://scialert.net/abstract/?doi=pjbs.1998.291.294
DOI: 10.3923/pjbs.1998.291.294
URL: https://scialert.net/abstract/?doi=pjbs.1998.291.294
INTRODUCTION
In view of the contribution of cotton, to the economy of Pakistan, the cotton plants has since long been subjected to extensive research. The efforts of the breeders were mainly aimed at, improving the genetic constitution of the plant affecting both its quantity and quality to be produced under wide range of environment of the country. The variability for the desirable traits in the existing germplasm is scanty and to generate variability, a number of methods are being used. Hybridization, backcrossing between two distant species having the same ploidy level and ovulo-embryo culture where the ploidy level is different, have been used to incorporate desirable genes into G. hirsutum. These methods involve the combining of whole genome of donor with the recipient and then to make selections in the segregating generations for desirable genotypes. It takes longer time to incorporate desirable gene(s) from the donor into the recipient genotype through these approaches. Moreover these methods are confronted with the problems due to, the differences in ploidy level, interspecific barriers and undesirable gene linkages present in the genotypes desired to be combined.
Induced mutaitons have been used to create genetic variability in many of the crop plants where the germplasm resources are scanty. In recent years a large number of early, high yielding, disease and insect resistance varieties of various crops, i.e. wheat, barley, rice, cotton etc. have been developed and released in different countries. Cotton variety NIAB 78 evolved through induced mutation at NIAB, institute is the good example in Pakistan. This approach involves the induction of mutations and selection of desirable mutants in segregating generations till uniformity is achieved. Therefore this method also requires longer time to obtain a new desirable and stable genotypes.
To overcome these problems, more recently various transformation approaches, like Agrobacterium mediated transfer of DNA, Electroporation method of gene transfer, particle bombardment method of gene transfer, DNA microinjection etc. are being used to incorporate desirable genes in lesser time. In cotton positive transformations (in vivo) are reported (Zhou et al., 1982,1983; Aslam et al., 1995, 1998), where the flower ovaries are injected with exogenous DNA. Agrobacterium mediated transformation is the most commonly used method for gene transfer in plants (Horsch et al., 1984). Although Agrobacterium has been used successfully for the transformation of a number of different plants species, difficulties exist due to limited host range, low efficiency of transformation, problems with the removal of Agrobacterium genes following transformation and manipulation of DNA in wide host range plasmids. Electroporation methods of gene transfer, using protoplast has also been used successfully for the production of transgenic plants (Fromm et al. 1985; Shillito et al., 1985). Some in vitro transformation methods being used are; DNA microinjections in tobacco (Crossway et al., 1986) in rye (De la Pena et al., 1987) and particle bombardment method in cotton (Finer and McMullen, 1990). However all these in vitro transformation methods require the use of callus culture to regenerate the whole plant from the transformed protoplast or cell. Moreover the ability of the cotton genotypes to regenerate from a cell into whole plant through callus tissue is genotype-specific (Trolinder and Xhixian, 1989). Genotype specificity of cotton has been further confirmed by Gawel and Robacker (1990). Since the cotton plant has shown genotype specificity towards callus formation. In vitro regeneration of the transformed cell into whole plant that expresses the inserted genes and that are fertile so that they can be passed on to the progeny is a major problem. Whereas DNA mediated embryo transformation is a straight forward approach and involves the injection of exogenous DNA solution into the plant reproductive structure under in vivo condition. The injected DNA solution transform the developing embryos during zygotic cell division and this transformation is heritable.
By adopting this approach useful genetic variability can be generated within a shorter period of time for the selection of desirable genotypes having few traits incorporated from the donor parent. Therefore the research studies were planned to incorporate, i) Cotton leaf curl virus (CLCV) disease resistance and white fly resistance from G. arboreum and ii) good fiber quality trait(s) from G. barbadense into G. hirsutum, through radiation and DNA-mediated embryo transformation technique.
MATERIALS AND METHODS
In these studies a locally cultivated tetraploid species of cotton; G. hirsutum (2n = 4x = 52) was used as recipient while the donor parents selected were i) a cultivated diploid species of cotton i.e. G. arboreum (2n = 2x = 26) and ii) a trtraploid long staple species, G. barbadense (2n = 4x = 52), but non-adaptive to local climate. Both the donor parents i.e. G. barbadense and G. arboreum are clearly distinguishable from the recipient; G. hirsutum in certain phenotypic and some economic traits. G. barbadense has longer, finer and stronger fibre as compared to G. hirsutum whereas G. arboreum carry resistance/immunity against insects and diseases, particularly cotton leaf curl virus (CLCV) disease. More than 200 plants of the recipient were raised from selfed seed in the field and at flowering 50 healthy plants were selected as experimental material. DNA from both the donors i.e. G. arboreum and G. barbadense was isolated using the protocol (Davis et al., 1986) as described below:
| i) | Seed of the donors were grown to obtain leaves for DNA extraction. |
| ii) | Approximately 5 gram of leaf tissue was pulverized in a mortar partially filled with liquid nitrogen. Finally grounded power was poured into 50 ml caped tube and liquid nitrogen was allowed to evaporate. Then 10 ml of 1xRDB buffer was added. |
| iii) | SDS (1 ml of 10% SDS) was added to pulverized tissue mixture. As a result the sample become viscous and DNA was released from the nuclei. |
| iv) | 11 mg proteinase K power (1 mg/1 ml concentration) was added and incubated for 1-2 hrs. at 37°C. Then another 11 mg of proteinase K was added and incubated for at least 1-2 hrs. until tissue was completely dissolved. |
| v) | 1 ml of 5 M NaCI was added. |
| vi) | Extract with an equal volume of SS phenol: chloroform: isomyl alcohol (10:10:1) was mixed gently but thoroughly by inverting the tube. Briefly spin to separate phases and upper aqueous phase was saved by transferring to a new tube. |
| vii) | Aqueous phase with two volumes of diethyl ether was extracted. |
| viii) | Aspirate off ether (on top) and saved lower aqueous phase. |
| ix) | 2.5 volumes of absolute ethanol was added to precipitate DNA. Large pieces of DNA were removed by collecting fibres with a glass rod and transferring them to a new tube. |
| x) | Dissolve pellet in 5 ml TE buffer overnight at 4°C. |
| xi) | To make the requisite solutions donors DNA was mixed, with protamine (1:1 ratio). |
| xii) | Half of the DNA solution from each donor was irradiated with gamma rays at 2.5 Gy and other half was used as such. |
The self-fertilized flower/ovaries of the recipient i.e. G. hirsutum were injected with donors (G. arboreum and G. barbadense) DNA solutions both irradiated and non-irradiated through the axil placenta, 24 hours post self pollination with a microsyringe. For each treatment 10 microliter solution was used and the injected flower/ovaries were protected from foreign contamination. Matured bolls from the treated flower/ovaries were collected, ginned and D0 generation seed was developed. D1 generation was raised from D0 seed at a spacing of 60×75 cm in the field. The D1 population was evaluated for phenotypic as well as economic traits changes incomparison to non-irradiated treatments and controls. The seed cotton from D1 population was harvested to develop D1 generation seed. D2 population was grown from D1 seed as plant progeny rows for evaluation and confirmation.
RESULTS AND DISCUSSION
Interploid interspecific studies: D0 generation seed obtained from interploid interspecific (G. hirsutum × G. arboreum DNA) injections with irradiated donor DNA and non-irradiated was studied in D1 population. The D1 population where the irradiated (2.5 Gy) donor DNA was used for injections exhibited higher percentage (15-20%) for plants resistance to cotton leaf curl virus (CLCV) disease. Whereas D1 population obtained from the treatments with non-irradiated donor DNA solutions had less percentage (10%) of plants resistance to CLCV disease (Table 1). Moreover the plants exhibiting resistance to CLCV disease had higher yield per plant and showed changes in boll size/weight, boll shape, plant type, hairiness and gossypol pigments and plant maturity. D1 generation plants and more hairiness, better boll size, more monopodial branches and good boll formation. However there were no changes for flower color and petal spot etc in D1 generation. Studies carried out on D2 generation indicated the persistency in the changes observed during D1 generation. However, some of the D2 progenies showed manifestation of qualitative traits i.e. petal spot, pollen and flower folour present in the donor G. arboreum. Moreover CLCV resistance progenies also showed segregation for CLCV resistance and CLCV susceptable genotypes. In this population 3:1 ratio was observed which indicated that CLCV gene behaved as monogenic and dominant (RR) trait (Aslam et al., 2000). Moreover the D2 generation progenies which showed resistance to CLCV disease also had higher yield than the standard and the recipient cotton genotypes (Table 2). The yield of the promising genotypes ranged from 2026.5 to 2602.2 (kg/ha) as compared to recipient (1020 kg/ha) and standard (620 kg/ha). Moreover the transformed genotypes seemed to be more tolerant to whitefly and further studies are in progress.
Intraploid interspecific studies: D0 generation seed obtained from non-irradiated interploid interspecific (G. hirsutum × G. barbadense DNA) injections with irradiated donor DNA was studied in D1 population. Phenotypic observations on various morphological traits showed changes for plant type, boll type, leaf size/shape, plant vigour etc. The plants expressing phenotypic changes had better boll weight, higher yield, fine, long and strong fibre as compared to recipient (Table 3). The results showed that the spectrum of changes for donor parents traits was higher where the irradiated (2.5 Gy) donor DNA was used for injections.
| Table 1: | Phenotypic and economic traits of D1 generation followed the injection of G. arboreum DNA into flower/ovaries of G. hirsutum |
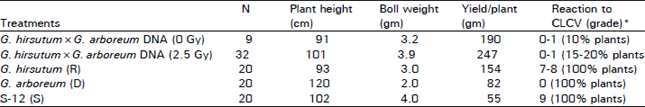 | |
| *Rating scale 0-9 i.e. 0 = Immune, 9 = Highly suseptible. R = Recipient, D = Donors, S = Standard | |
| Table 2: | Characteristics of the promising plant progenies of G. hirsutum×G. arboreum DNA injections in D2 generation |
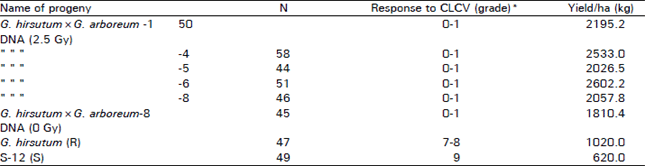 | |
| *Rating scale 0-9 i.e. 0 = Immune, 9 = Highly susceptible. R = Recipient, S = Standard = G. hirsutum Note: Some progenies had petal spot and yellow pollen colour etc. | |
| Table 3: | Phenotypic and economic traits (Range) of D1 generation followed injections of G. barbadense into flower ovaries of G. hirsutum |
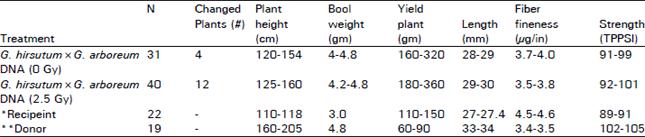 | |
*G. hirsutum, **G. barbadense. Note: The changed plants: i) Were more faster in growth; ii) Had larger leaves conical bolls and long petioles; iii) Had larger to medium flower with bigger calyx iv) Had no evidence of marker genes | |
| Table 4: | Characteristics of the promising transformed progenies of G. hirsutum×G. arboreum DNA injections in D2 generation |
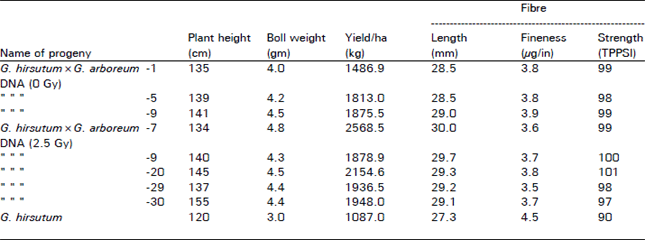 | |
| Note: Besides other D1 phenotypic changes the D2 plant also had transformation for i) Red flower petal spot, ii) Yellow flower colour, iii) and Yellow pollen colour | |
The results obtained on D2 generation plant progeny rows showed consistency in the changes noted during D1 generation. Morever some of the D2 generation progenies expressed changes in the qualitative traits i.e. red petal spot, pollen colour and flower colour etc. like the donor parent G. barbadense (Table 4). The transformed progenies had higher boll weight (4-4.8 gram) than the recipient (3 gram). The yield of the transformed progenies ranged from 1486.9 to 2568.5 kg/ha as compared to 1087 kg/ha of the recipient. Moreover the transformed progenies had better fibre finess i.e. 3.9 to 3.6 μg/in. as compared to 4.5 μg/in of the recipient. The fibre strength of 101 TPPSI was obtained. These progenies also had better staple length than the recipient.
The results of D1 and D2 generation obtained from the injection with donors (G. arboreum and G. barbadense) DNA irradiated and as such into G. hirsutum flower/ovaries 24 hours post self pollination revealed the incorporation and preservation of the donor parents traits ino the recipient. Moreover the frequency of the donor gene incorporation increased where the donors DNA was irradiated at low doses (2.5 Gy) of gamma rays before injections (Kohler et al., 1989). The research studies illustrated higher transformations for boll size/weight, fibre fineness and confirmed the earlier studies reported by Aslam et al. (1995, 1998).
ACKNOWLEDGMENTS
Acknowledgments are due to international Atomic Energy Agency (IAEA) for providing financial support for these studies under IAEA research contract 8142/RB.
REFERENCES
- Aslam, M., M.T. Elahi and N. Iqbal, 1998. Introgression of G. barbadense genes into G. hirsutum through DNA-mediated embryo transformation approach. Pak. J. Biol. Sci., 1: 11-14.
CrossRefDirect Link - Aslam, M., C. Jiang, R. Wright and A.H. Paterson, 2000. Identification of molecular markers linked to leaf curl virus disease resistance in cotton. J. Sci. I. R. Iran, 11: 277-280.
Direct Link - Crossway, A., J.W. Oakes, J.M. Irvine, B. Ward, V.C. Knauf and C.K. Shewmaker, 1986. Integration of foreign DNA following microinjection of tobacco mesophyll protoplasts. Mol. Gen. Genet., 202: 179-185.
CrossRefDirect Link - De la Pena, A., H. Lorz and J. Schell, 1987. Transgenic rye plants obtained by injecting DNA into young floral tillers. Nature, 325: 274-276.
CrossRefDirect Link - Finer, J.J. and M.D. McMullen, 1990. Transformation of cotton (Gossypium hirsutum L.) via particle bombardment. Plant Cell Rep., 8: 586-589.
CrossRefDirect Link - Fromm, M., L. Taylor and V. Walbot, 1985. Expression of genes transferred into monocot and dicot plant cells by electroporation. Proc. Natl. Acad. Sci. USA., 82: 5824-5828.
Direct Link - Gawel, N.J. and C.D. Robacker, 1990. Genetic control of somatic embryogenesis in cotton petiole callus cultures. Euphytica, 49: 249-253.
Direct Link - Horsch, R.B., J.E. Fry, N.L. Hoffman, D. Eichholtz, S.A. Rogers and R.T. Fraley, 1985. A simple and general method for transferring genes into plants. Science, 227: 1229-1232.
Direct Link - Kohler, F., G. Cardon, M. Pohlman, R. Gill and O. Schieder, 1989. Enhancement of transformation rates in higher plants by low-dose irradiation: Are DNA repair systems involved in the incorporation of exogenous DNA into the plant genome? Plant Mol. Biol., 12: 189-199.
CrossRefDirect Link - Shillito, R.D., M.W. Saul, J. Paszkowski, M. Muller and I. Potrykus, 1985. High efficiency direct gene transfer to plants. Nat. Biotechnol., 3: 1099-1103.
CrossRefDirect Link - Trolinder, N.L. and C. Xhixian, 1989. Genotype specificity of the somatic embryogenesis response in cotton. Plant Cell Rep., 8: 133-136.
CrossRefDirect Link - Zhou, G., J. Weng, Y. Zeng, J. Huang, S. Qian and G. Liu, 1983. Introduction of exogenous DNA into cotton embryos. Methods Enzymol., 101: 433-481.
PubMedDirect Link