
M. Ahsan
School of Agriculture and Forest Science, University of Wales, Bangor, Gwyndd LL57 2UW, UK
D. Wright
School of Agriculture and Forest Science, University of Wales, Bangor, Gwyndd LL57 2UW, UK
Pakistan Journal of Biological Sciences
Year: 1998 | Volume: 1 | Issue: 3 | Page No.: 209-211







ABSTRACT
Effects of leaf age on ion uptake (Na+, K+ and CI‾) and K+/Na+ ratio were investigated using the three S1 wheat selections Alex-1, KRL-24 and Khar-1. Na+ and CI‾ contents were found to be increasing in the older leaves, but K+ content and K+/Na+ ratio were found to be decreasing in the younger leaves. However, it is suggested from the results that sampling should be done when leaves are fully expanded. Khar-1 was found to be intermediate between Alex-1 and KRL-24 in ion uptake and K+/Na+ ratio at three out of the four sampling dates. The considerable variation in Na+, K+, Cl‾ uptake and K+/Na+ ratio between sampling dates suggests that these traits might be less useful as selection criteria.
PDF Abstract XML References Citation
How to cite this article
M. Ahsan and D. Wright, 1998. Effects of Leaf Age on Ion Uptake in Wheat (Triticum aestivum L.) Under Saline Conditions. Pakistan Journal of Biological Sciences, 1: 209-211.
DOI: 10.3923/pjbs.1998.209.211
URL: https://scialert.net/abstract/?doi=pjbs.1998.209.211
DOI: 10.3923/pjbs.1998.209.211
URL: https://scialert.net/abstract/?doi=pjbs.1998.209.211
INTRODUCTION
It is considered by many research workers that biological variation in Na+ and K+ contents is an important factor in the genetic basis of salt tolerance in wheat (Joshi et al., 1979; Shah et al., 1987; Singh et al., 1988; Gorham, 1988). However, ion content can be used as breeding tool for selecting salt tolerant genotypes. Maas and Grieve (1994) found that response of crop plant to salt stress at different growth stages is different. Cramer et al. (1994) reported that relative growth rate (RGR) and leaf area ratio (LAR) were inhibited by salinity in the early stages of stress. Therefore it is very important to know the extent of variation in Na+, K+ and Cl‾ contents in plant leaves. Many research workers have measured ion contents, usually by sampling at a single time from fully expended fourth or flag leaves. However it is very important to know the pattern of ion uptake of genotypes, because genotypes initially with a low content might have a higher content at later growth stages. In breeding crop plants, differences in phenology are also important. When comparing early and late maturing varieties it is impossible to harvest the same leaf from all plants, at the same growth stage and on the same day. Such variation could give misleading information if differences in maturity are significantly large.
Jones and Qualset (1984) suggested that precise and efficient analytical techniques are needed to confirm such biological variation in plants. However, differences in leaf age must be identified because it is important in determining the ionic differences between tolerant and sensitive wheat genotypes. The experiment reported here were done to examine if differences between genotypes in Na+, K+, Cl‾ contents and K+/Na+ ratio were consistent over a range of sampling dates.
MATERIALS AND METHODS
This experiment was conducted in glass-house at Pen Y Ffridd Field Station, University of Wales, Bangor, UK., during October to December, 1994. The temperature of the glass-house was maintained at 18-20°C. The natural day light was supplemented when necessary by 400 W Son-T Sodium vapour lamps to provide a photoperiod of 16 hrs. Three S1 wheat selections Alex-1, KRL-24 and Khar-1 were selected from within three wheat varieties Alexandria (salt-sensitive), KRL1-4 (salt-tolerant) and Kharchia-65 (salt-tolerant) on the bases of high K+/Na+ ratio (Ahsan, 1996). These three S1 selections were tested at 100 mol m–3 NaCI. The seeds were grown in the glass-house on capillary matting starting on 24-10-1994. Seedlings were transplanted into hydroponic culture on 3-11-1994. A total of 60 plants (20 plants per selection per replication) were grown in three replications. A completely randomized design was used. The size of pot was 52x35x16 cm. The pots were well aerated. The plant-to-plant and row-to-row distance was 3.5 and 6.0 cm respectively. Salt stress (100 mol m–3 NaCI) was introduced in three increments over a period of five days starting on 9-11-1994. Phostrogen (0.5 g l–1 phostrogen Ltd., Corwen, Clwyd, UK) was applied to each pot. Phostrogen is blended 10-10-27 NPK fertilizer with 1.3 percent Mg, 0.4 percent Fe and 0.02 percent Mn. A modified Long Ashton Solution (Hewitt, 1966) was used in combination with phostrogen to supply micro-nutrients. Fourth leaves from two plants per S1 selection per replication were sampled, 23, 30, 37 and 44 days after transplanting. The leaves were rinsed quickly in distilled water and blotted dry with tissue paper. The samples were placed in Eppendorf tubes and stored in a freezer set at -10̊C. Cell sap was extracted by following the method (Gorham et al., 1984). The cell sap was diluted with distilled water. Na+ and K+ contents were estimated form diluted cell sap by using the atomic absorption spectrophotometer (Model-151, Instrumentation Laboratory) and K+/Na+ ratio was determined. Chlorides were measured with an ion selective electrode (Microprocessor lonalyzer/109). Statistical analysis were performed by using the Minitab, SYSTAT statistical packages. Analysis of variance (ANOVA) were used to assess significant differences (p<0.05) between means of the S1 wheat selections.
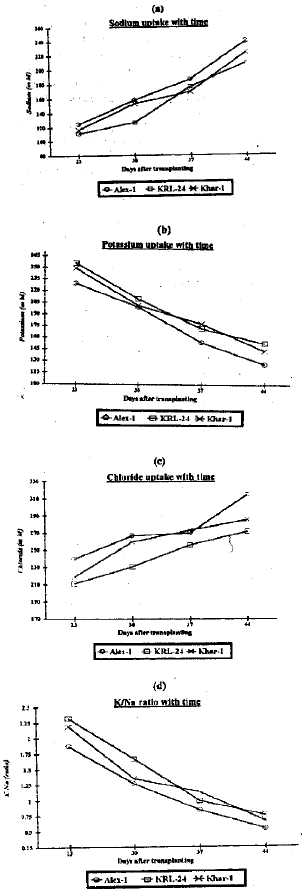 | |
| Fig. 1: | Effects of leaf age on (a) Na+ content (mol m–3), (b) K+ content (mol m–3), (c) Cl‾ content (mol m–3) and (d) K+/Na+ (ratio) in S1 selections from within three wheat varieties under saline conditions. |
RESULTS AND DISCUSSION
The trend in all S1 lines was for Na+ and Cl‾ contents to increase and for K+ content and K+/Na+ ratio to decrease with leaf age. Similarly, Begum et al. (1992) reported in wheat that salt stress increased Na+ and Cl‾ accumulation, while it decreased K+ accumulation. The tends in Na+, K+, CI‾ contents and Kl+/Na+ ratio were found consistent between Alex-1 and KRL-24. Although Khar-1 was found to be less consistent than Alex-1 and KRL-24 (Fig. 1). It is concluded from these results that leaves should be sampled when they are fully expanded.
Most of the differences were non significant between S1 selections for Na+, CI‾, K+ contents and K+/Na+ ratio, except that KRL-24 had significantly (p<0.05) higher K+/Na+ ratio than Alex-1 at 30 days after transplanting. It is possible that, all these S1 selections were selected on the basis of high K+/Na+ ratio. Alex-1 also had significantly (p<0.05 ) higher Cl‾ content than KRL-24 at 23 and 30 days after transplanting (Ahsan, 1996).
The differences between S1 selection in ion content and K+/Na+ ratio were found less consistent in this study. Khar-1 was found to be intermediate between Alex-1 and KRL-24 at three out of the four sampling dates (Fig. 1), However the considerable variation in Na+, K+, Cl‾ contents and K+/Na+ ratio with leaf age suggests that these physiological traits may be less useful while comparing genotypes. Various research workers evaluated different characters for their potential as selection criteria for salt tolerance. For early secreening of wheat genotypes, germinability at high salt concentration (Roy, 1991) and seedling dry and fresh weight at different levels of salinity (Prakash and Sastry, 1992) along with Na+ and K+ contents are useful criterion for salt tolerance. Leaf position in two different growing systems (soil and hydroponic culture) also indicated considerable biological variation in ion uptake. Most of the linear, correlation between the fourth and flag leaf in ion uptake were found significant in soil and non-significant in the hydroponic culture supports the idea that these physiological traits are less useful while using as selection criteria in crop plants (Ahsan, 1996). Ashraf and McNeilly (1988) proposed a general selection criterion for salt tolerance as they suggested the use of whole plant performance for assessment of salt tolerance of wheat.
REFERENCES
- Ashraf, M. and T. McNeilly, 1988. Variability in salt tolerance of nine spring wheat cultivars. J. Agron. Crop Sci., 160: 14-21.
CrossRefDirect Link - Begum, F., J.L. Karmoker, Q.A. Fattah and A.F.M. Muniruzzaman, 1992. The effect of salinity on germination and its correlation with K+, Na+, Cl-accumulation in germinating seeds of Triticum aestivum L. cv. Akbar. Plant Cell Physiol., 33: 1009-1014.
Direct Link - Cramer, G.R., G.J. Alberico and C. Schmidt, 1994. Leaf expansion limits dry matter accumulation of salt-stressed maize. Aust. J. Plant Physiol., 21: 663-674.
Direct Link - Gorham, J., E. McDonnell and R.G.W. Jones, 1984. Pinitol and other solutes in salt-stressed Sesbania aculeata. Zeitschrift Pflanzenphysiologie, 114: 173-178.
CrossRefDirect Link - Jones, R.A. and C.O. Qualset, 1984. Breeding Crops for Environmental Stress Tolerance. In: Application of Genetic Engineering to Crop Improvement, Collins, G.B. and J.B. Petolino (Eds.). Martinus Nijhoff/Dr. W. Junk Publishers, The Netherlands, ISBN: 978-94-009-6209-5, pp: 305-340.
Direct Link - Joshi, Y.C., A. Qadar and R.S. Rana, 1979. Differential sodium and potassium accumulation related to sodicity tolerance in wheat. Indian J. Plant Physiol., 22: 226-230.
Direct Link - Shah, S.H., J. Gorham, B.P. Forster and R.G. Wyn Jones, 1987. Salt tolerance in the triticeae: The contribution of the d genome to cation selectivity in hexaploid wheat. J. Exp. Bot., 38: 254-269.
CrossRefDirect Link - Singh, K.N., S.K. Sharma and K.N. Singh, 1988. Promising new wheat varieties for salt-affected soils. Indian Farm., 38: 21-25.
Direct Link