
R. Haritha
Department of Biotechnology, College of Science and Technology, Andhra University, Visakhapatnam 530003, India
K. SivaKumar
Department of Biotechnology, College of Science and Technology, Andhra University, Visakhapatnam 530003, India
A. Swathi
Department of Biotechnology, College of Science and Technology, Andhra University, Visakhapatnam 530003, India
Y.S.Y.V. Jagan Mohan
Department of Biotechnology, College of Science and Technology, Andhra University, Visakhapatnam 530003, India
T. Ramana
Department of Biotechnology, College of Science and Technology, Andhra University, Visakhapatnam 530003, India







ABSTRACT
A black pigmented, proteolytic actinomycete strain was isolated from a marine sediment sample collected near Kakinada of Andhra Pradesh province, India. The isolate grows to produce circular colonies that had yellow-white substrate mycelium and grey aerial mycelium on Starch Casein Agar, with black diffusible pigment. Its proteolytic activity was indicated by growth and clear zone appearance on casein-skim milk agar media plate. Phylogenetic analysis based on 16S rRNA gene sequences showed that the strain belongs to the genus Streptomyces, with the highest similarity to Streptomyces carpaticus NBRC15390T (99.6%). On the basis of polyphasic evidence, the strain was recognized as a new isolate of Streptomyces carpaticus from marine source. Optimal conditions for protease production by Streptomyces carpaticus were; incubation temperature of 35°C, pH 7.0, inoculum size 6% (v/v) and agitation speed of 150 rpm. The best carbon, organic nitrogen and inorganic nitrogen sources for production of protease enzyme were glucose, soybean meal and ammonium sulfate, respectively. Under these conditions the newly isolated strain produced protease activity of 17542.51 U mL-1. This is the first report on protease production by Streptomyces carpaticus from marine source.
PDF Abstract XML References Citation
Received: August 03, 2011;
Accepted: November 23, 2011;
Published: January 21, 2012
How to cite this article
R. Haritha, K. SivaKumar, A. Swathi, Y.S.Y.V. Jagan Mohan and T. Ramana, 2012. Characterization of Marine Streptomyces carpaticus and Optimization of Conditions for Production of Extracellular Protease. Microbiology Journal, 2: 23-35.
DOI: 10.3923/mj.2012.23.35
URL: https://scialert.net/abstract/?doi=mj.2012.23.35
DOI: 10.3923/mj.2012.23.35
URL: https://scialert.net/abstract/?doi=mj.2012.23.35
INTRODUCTION
The Streptomyces have offered over 50 years of interest to industry and academia (Anderson and Wellington, 2001; Berdy, 2005), for their unrivaled ability to elaborate different kinds of secondary metabolite compounds with a range of biological activities that may ultimately find application as anti-infectives, anti-cancer agents or other pharmaceutically useful compounds (Kim and Hwang, 2003; Bibb, 2005). The marine Streptomyces species are able to produce a variety of bioactive compounds, including extracellular enzymes (El-Dein et al., 2010; El-Sersy et al., 2010; El-Shafei et al., 2010). By virtue of their unique natural habitat, the enzymes from marine microorganisms may carry novel physiochemical properties which can be desirable features from a biotechnological perspective.
Proteases represent one of the three largest groups of industrial enzymes and have traditionally held the predominant share of the industrial enzyme market accounting for about 60% of total worldwide sale of enzymes (Singh et al., 2001; Chu, 2007; Oskouie et al., 2008). The immense diversity of proteases, in contrast to the specificity of their action has attracted worldwide attention to exploit their biotechnological and industrial applications, such as laundry detergents, leather preparation, food industry and protein recovery or solubilization, organic synthesis, silver recovery from photographic film, digestive aids and anti-inflammatory agents (Chen et al., 2004; Kasana and Yadav, 2007). The great economic value of proteases still gives an impulsion to the search for new proteases with novel properties.
In this report, we describe the taxonomic characterization of a potent protease producing marine strain, as a new isolate of Streptomyces carpaticus. This is the first report of S. carpaticus from a marine source. We also describe a method to improve protease production by optimizing medium composition.
Isolation of marine actinomycetes: The marine sediment near Kakinada [Latitude (N) 16°59.507’; Longitude (E) 82°43.923’] coastal region, the south east coast of India was collected at a depth of 108.05 cm. Actinomycetes were isolated by the standard serial dilution technique (Jensen et al., 1991) using starch casein agar media (Kuster and Williams, 1964) and the plates were incubated at 28°C for 7-45 days. The actinomycetes isolates obtained were purified and maintained on the starch casein agar medium as a working culture.
Screening of marine actinomycete isolates for proteolytic activity: A qualitative screening for the proteolytic activity of the isolates was indicated by growth and clear zone appearance on casein-skim milk agar media plates (Williams and Cross, 1971).
Taxonomic identification of actinomycete isolate: Polyphasic taxonomic approach was used to identify the active marine isolate. Morphological, cultural, physiological and biochemical characterization of the strain were studied by following the methods of Shirling and Gottlieb (1966). The morphological characteristics of strain BTSS-501T were assessed by scanning electron microscopy (model JSM-6610 LV; JEOL, Ltd., USA) of 14 day-old cultures on ISP 2 medium. A range of physiological and biochemical characteristics were examined according to the standard protocols of Goodfellow (1971) and Williams et al. (1983). Standard analytical procedures were used to extract and analyze the isomeric forms of diaminopimelic acid (Hasegawa et al., 1983), whole-organism sugars (Staneck and Roberts, 1974), isoprenoidquinones (Minnikin et al., 1984; Collins, 1985), polar lipids (Minnikin et al., 1984) and fatty acids (Sutcliffe, 2000). The phylogenetic trees of the almost complete 16S rRNA sequence and the variable γ region (positions 158-277) of 16S rRNA gene sequence of the strain were constructed using the maximum-likelihood (Felsenstein 1981) tree-making algorithm from the PHYLIP software package. The G+C content of the genomic DNA was determined spectrophotometrically using the thermal denaturation method (Marmur and Doty, 1962).
This strain was deposited in Microbial Type Culture Collection and Gene Bank, India (MTCC) as MTCC 33991T.
Culture conditions for enzyme production: One hundred milliliter of the production medium was inoculated with 1 mL of overnight culture of protease-producing isolate and was incubated in orbital shaker at 28°C for 96 h in 250 mL conical flasks. The cell free supernatant of the fermented broth obtained by the centrifugation at 10,000 rpm for 10 min at 4°C was used as the crude enzyme source. The production media contained (g L-1) soluble starch 20, yeast extract 5, peptone 5, glucose 5, casein 5, soyabean meal 5, (NH4)2SO4 6, NaCl 5, CaC03 6 and pH 7.0.
Quantitative assay of protease: Protease activity in the culture filtrate was measured according to the method of Keay and Wildi (1970) method. One unit (U) of protease activity is equivalent to μg of tyrosine equivalent liberated per mL of enzyme at 37°C for 30 min.
Comparison of protease production by Streptomyces carpaticus with reference bacterial strain, Bacillus cereus MTCC 430: The protease producing reference strain, Bacillus cereus MTCC 430 was obtained from the Microbial Type Culture Collection, Institute of Microbial Technology, Chandigarh, India. Both, B. cereus MTCC 430 and S. carpaticus were grown separately in 50 mL of production medium in 250 mL flasks at 28°C for 7 days at 120 rpm. At every 2 days intervals, the cultures were harvested and the cell free culture supernatants of both the organisms were prepared as mentioned above. The protease activity in the culture supernatants was determined by quantitative enzyme assay as described above.
Optimization of protease production: The effect of nutrients on growth and protease production was tested by adding different carbon sources (1%, w/w) and inorganic nitrogen sources at 0.1% (w/v) level and organic nitrogen sources at 0.05% (w/v) level into the Pridham and Gottlieb’s inorganic salts medium (Pridham and Gottlieb, 1948). The effect of experimental conditions on protease production including incubation temperatures (4-60°C), agitation conditions (30-240 rpm), inoculum sizes and initial pH (4.0-11.0) were investigated. All the experiments were done in triplicates.
Nucleotide sequence accession number: The 16S rRNA gene sequence of the new strain determined in this study has been deposited in GenBank under the accession number HQ711933.
RESULTS AND DISCUSSION
Isolation and Screening of marine actinomycete isolates for proteolytic activity: A total of 5 different actinomycetes were isolated from marine sample collected from the Bay of Bengal, India. Qualitative assay of protease revealed that amid the 5 marine actinomycetes, 4 isolates produced protease. Among the 4, an active isolate was selected for further studies as it produced significant clear proteolytic zone (Fig. 1).
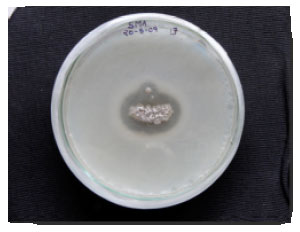 | |
| Fig. 1: | Proteolytic activity of the marine isolate on casein-skim milk agar |
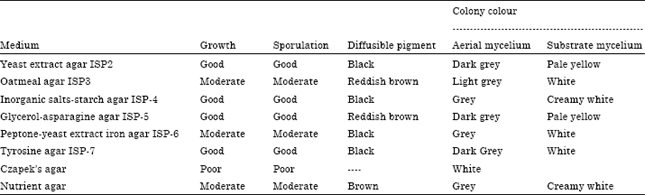
Taxonomic identification of actinomycete isolate BTSS-501T: The active strain was Gram-positive, non-acid-fast, nonmotile, filamentous actinomycete. Morphological observations of the culture of the strain on different ISP media are presented in Table 1. Substrate hyphae of the strain were extensively branched and long, spiral spore chains were borne on the aerial hyphae. The oval spores (dimensions 0.5-0.7x0.6 μm) were non-motile with smooth spore surface (Fig. 2). Detailed results of the physiological characterization and a summary of the differentiating physiological characters of the strain and the most closely related species is shown in Table 2. Chemotaxonomic tests showed that the cell wall contained LL-diaminopimelic acid and traces of glycine. No diagnostic sugars were detected in whole-cell hydrolysates. The predominant menaquinones were MK-9 (H4)-69.7%, MK-9 (H6)-14.1% and MK-9 (H8)-16.2% and the diagnostic phospholipids were phosphatidylethanolamine and phosphatidylcholine. Fatty acids comprised iso-C14:0 (5.92%), iso-C15:0 (3.00%), anteiso-C15:0 (3.67%), C16:0 iso H (3.51%), iso-C16:0 (49.11%), C16:0 (16.65%), anteiso-C17:0 (2.70%), C17:1ω8c (1.65%) and C17:0 cyclo (4.74%). In addition, the sum of C16:1ω7c/C15:0 iso 2-OH was 9.06%. The DNA G + C content was 72.5%.
| Table 1: | Cultural characteristics of the strain on various ISP media |
 | |
 | |
| Fig. 2: | Scanning electron micrograph indicates the spore chain morphology of marine actinobacteria strain grown on ISP medium 2 for 3 weeks at 28°C. Bar 2 μm |
| Table 2: | Differentiating physiological characteristics of the new strain of actinomycetes and related species |
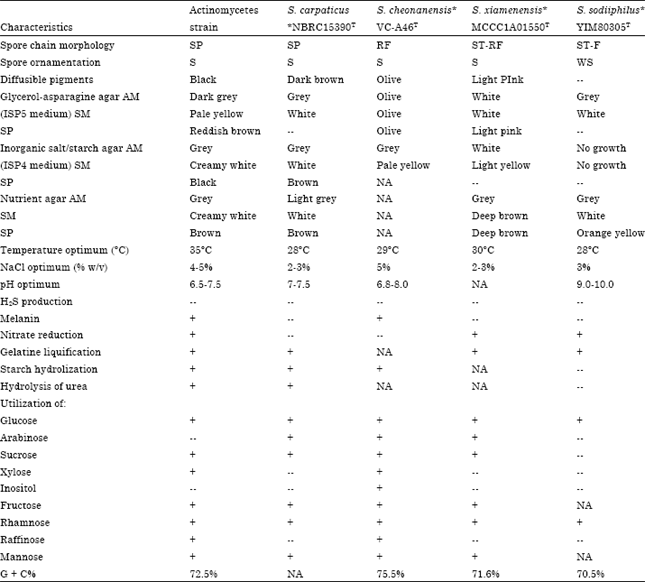 | |
| aAbbreviations: AM: Aerial mycelium, SM: Substrate mycelium, SP: Soluble pigment, ST: Straight, RF: rectiflexibiles, SP: Spirales, F, flexuous, WS: Wrinkled surface, S: Smooth. b Symbols: +, utilization;--, not utilization; NA, Data not available. *Data for reference type species strains were taken from Li et al. (2005), Hyo et al. (2006) and Xu et al. (2009) | |
The variable γ region sequences (158-277 nt) of the 16S rRNA gene, indicated that the marine actinobacteria strain was clustered with Streptomyces cheonanensis VC-A46T, Streptomyces sp. A452 Ydz-DST and Streptomyces sp. C12 (2006)T (data not shown). A BLAST search of the GenBank database using the 1471 bp 16S rRNA gene sequence of strain BTSS-501T showed its similarity to that of many species of the genus Streptomyces. The 16S rRNA gene sequence of marine actinobacteria strain showed levels of homology of 99.6% (5 nucleotide differences out of 1,455) to S. carpaticus NBRC15390T (GenBank accession No. AB184641), 99.5% (6 nucleotide differences out of 1,466) to S. cheonanensis VC-A46T (GenBank accession No. AY822606), 97.4% (38 nucleotide differences out of 1,470) to Streptomyces xiamenensis MCCC1A01550T (GenBank accession No. EF012099) and 97.1% (42 nucleotide differences out of 1,469) to Streptomyces sodiiphilus YIM80305T (GenBank accession No. AY236339).
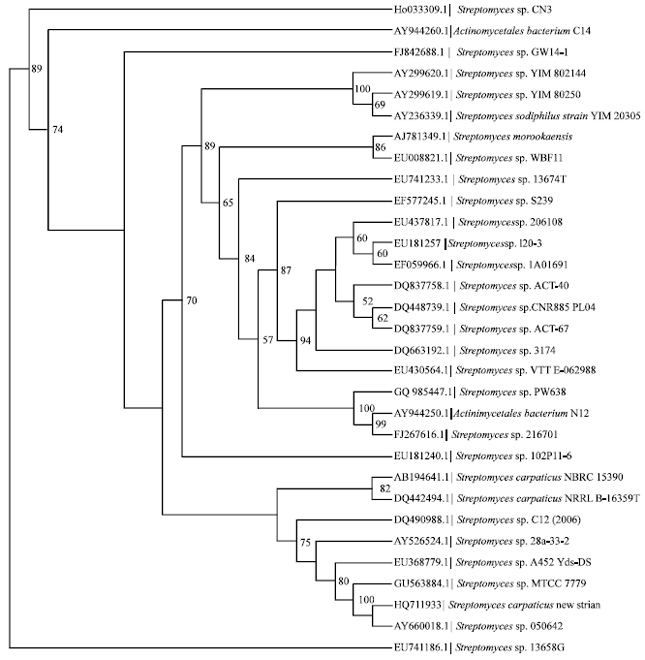 | |
| Fig. 3: | Phylogenetic maximum-likelihood tree based on 16S rRNA gene sequences showing the relationship between marine active strain and related members of the genus Streptomyces. Streptomyces niger strain AS 4.1244 (Accession no.FJ406109.1) was used as an outgroup. Bootstrap values (expressed as percentages of 1000 replications) greater than 50% are given at nodes. Bar 0.01 substitutions per nucleotide positions |
The phylogenetic tree indicated that this strain formed a distinct branch with an undescribed isolate, Streptomyces sp., 050642 (Hong, K. and Yan, L, unpublished; GenBank accession No. AY660018.1), derived from near-shore sediments of Hainan Island, China, within the S. carpaticus NBRC15390T (the top BLAST result) clade (Fig. 3). Phylogenetic analysis based on 16S rRNA gene sequences revealed that strain should be assigned to the genus Streptomyces.
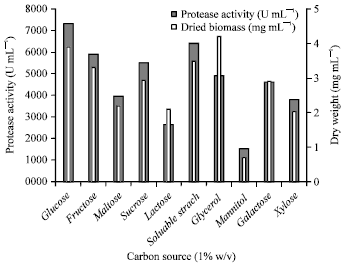 | |
| Fig. 4: | Effect of carbon sources on protease production and growth of Streptomyces carpaticus |
Based on the phenotypic, phylogenetic and chemotaxonomic analysis, it is evident that the isolated strain is different from previously described S. carpaticus. Therefore, this strain represents a new isolate of S. carpaticus from marine source.
Quantitative comparison of protease production by S. carpaticus with reference bacterial strain, Bacillus cereus MTCC 430: Result of quantitative assay of protease has shown that S. carpaticus produced high protease activity of 6326.8 Units mL-1 on 4th day compared to reference bacterial strain, B. cereus MTCC 430 which produced 2835.71 Units mL-1 on 3rd day (data not shown). The results clearly illustrate the hyper protease producing potential of marine S. carpaticus compared to the already known protease producing bacterial strain. Hence, this strain is useful for large scale protease production.
Effect of nutritional factors on protease production
Effect of carbon sources on the production of protease: The results presented in (Fig. 4) demonstrate that Glucose was the best carbon source for protease production (7310.29 U mL-1) while Mannitol and Lactose caused 78.93 and 63.84% decrease in protease yield. Glycerol incorporated medium showed the maximum cell growth. Present results are in accordance with Shafee et al. (2005) and Bhatiya and Jadeja (2010) who reported that, the protease production reached to the maximum when glucose was added to the medium and on the contrary to previous investigations which showed that glucose caused repression of protease production (Sen and Satyanarayana, 1993; Benslimane et al., 1995).
Effect of various nitrogen sources on protease production: Protease production was detected in medium containing inorganic nitrogen sources (Fig. 5). Ammonium sulphate gave the highest protease production (8731.49 U mL-1) and cell growth. This is in accordance with the results of El-Safey and Abdul-Raouf (2004) and Seri Intan et al. (2005) who observed that, optimum inorganic nitrogen source for protease production was (NH4)2SO4 by Bacillus subtilis and Bacillus pumilis, respectively. Supplementation of medium with sodium nitrate and potassium nitrate reduced the protease yield by 71.3 and 67%, respectively. The cell growth values obtained with inorganic nitrogen sources such as sodium nitrate and potassium nitrate were considerably lower than those observed in ammonium sulphate-incorporated medium.
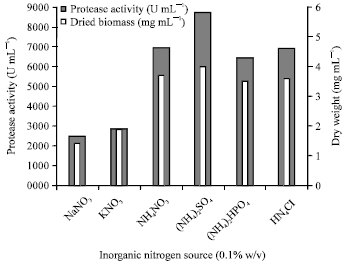 | |
| Fig. 5: | Effect of inorganic nitrogen sources on protease production and growth of Streptomyces carpaticus |
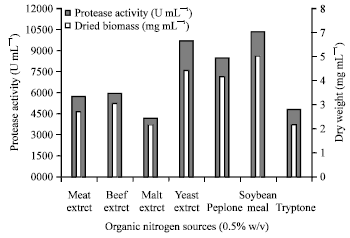 | |
| Fig. 6: | Effect of organic nitrogen sources on protease production and growth of Streptomyces carpaticus |
The ability of Streptomyces carpaticus to produce protease in liquid media was examined in various organic nitrogen sources (Fig. 6). Soybean meal exihibited the highest level of cell growth and protease production (10379.42 U mL-1). Similarly, soybean meal was reported as the best nitrogen source for Streptomyces avermectinus (Sen and Satyanarayana, 1993), Bacillus licheniformis (Ahmad et al., 2008) and mesophilic bacterium (Bhatiya and Jadeja, 2010). Malt extract and Tryptone were ineffective as nitrogen sources to promote the growth and protease production of the strain.
Effect of experimental conditions on protease production
Effect of temperatures on protease production: The optimum temperature for growth and protease production by the strain was 35°C (11589.09 U mL-1). Higher temperatures at 40 and 45°C caused more than 42.1 and 59.63% reduction in enzyme production at 96 h incubation (Fig. 7). No actinobacterial growth and protease production were observed at 4 or at 55°C and above temperatures.
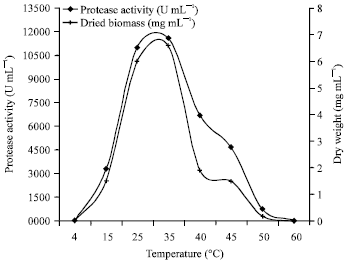 | |
| Fig. 7: | Effect of temperature on protease production and growth of Streptomyces carpaticus |
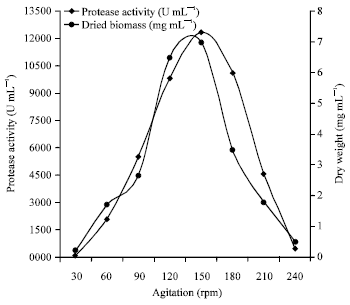 | |
| Fig. 8: | Effect of agitation rates on protease production and growth of Streptomyces carpaticus |
These results confirmed the observation of Qadar et al. (2009) who reported that the optimum temperature for protease production by Bacillus sp., is 35°C. Hence, based on the optimum temperature, Streptomyces carpaticus was classified as mesophile.
Effect of agitation rates on the protease production: Agitation is another important parameter that effects enzyme production in bacteria. The results represented in Fig. 8 revealed that enzyme production increases with increasing speed. The maximum protease production was observed at 150 rpm (12318.27 U mL-1). However, enzyme production was reduced by further increases in agitation rate. This finding was consistent with that of Darah and Ibrahim (1996) who reported that, high agitation speed caused a drop in enzyme production due to excessive breakdown of the enzyme or cell lysis or excessive cell permeability related to abrasion by shear forces.
Effect of inoculum sizes on the protease production: The result revealed that, protease production and actinobacterial growth (Fig. 9) were optimum when 6.0% (v/v) of inoculum was used (15390.48 U mL-1).
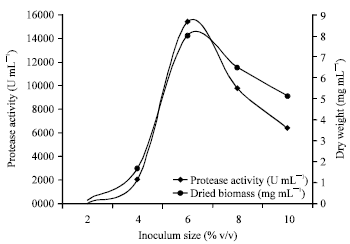 | |
| Fig. 9: | Effect of inoculums sizes on protease production and growth of Streptomyces carpaticus |
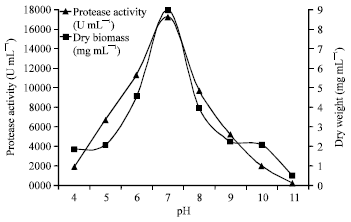 | |
| Fig. 10: | Effect of pH on protease production and growth of Streptomyces carpaticus |
Higher inoculum size at 8.0% (v/v) and 10.0% (v/v) decreased the protease production by 36.3 and 58.4%, respectively. Therefore, high inoculum sizes might not necessarily give higher protease yield or cell growth alternatively higher inoculum sizes could result in the lack of oxygen and nutrient depletion in the culture media.
Effect of pH on the protease production: In this study neutral pH 7.0 maximized protease (17542.51 U mL-1) and cell growth production (Fig. 10). At acidic pH (pH 6.0), 34.0% reduction of the enzyme production was observed compared to pH 7.0. At pH 8.0, the protease yield was decreased by 43.0% after 96 h incubation compared to pH 7.0. The optimum pH for protease production of Streptomyces carpaticus was similar to that reported for Streptomyces fradiae (Ellaiah and Srinivasulu, 1996) and Streptomyces roseiscleroticus (Vonothini et al., 2008).
CONCLUSION
This is the first report of marine Streptomyces carpaticus producing protease. Considering the high activity, it is proposed that the extracellular protease produced by marine Streptomyces carpaticus holds enormous scope for commercial exploitation. In this context, optimum conditions for production of protease were investigated. There is abundant scope for further research on enzyme characterization, structure elucidation and enzyme engineering of this protease towards development of the enzyme for a wide array of applications.
ACKNOWLEDGMENT
This study was supported by the University Grants Commission, New Delhi, India. We are grateful to Department of Science and Technology-Promotion of University Research for Scientific Excellence (DST-PURSE) programme of Andhra University for providing JSM-6610 LV; JEOL model Scanning Electron Microscope.
REFERENCES
- Ahmad, S.A., R.A. Al-Domany, N.M.A. El-Shayeb, H.H. Radwan and S.A. Saleh, 2008. Optimization, immobilization of extracellular alkaline protease and characterization of its enzymatic properties. Res. J. Agric. Biol. Sci., 4: 434-446.
Direct Link - Berdy, J., 2005. Bioactive microbial metabolites: A personal view. J. Antibiot., 58: 1-26.
CrossRefDirect Link - Bibb, M.J., 2005. Regulation of secondary metabolism in streptomycetes. Curr. Opin. Microbiol., 8: 208-215.
CrossRefPubMedDirect Link - Chu, W.H., 2007. Optimization of extracellular alkaline protease production from species of Bacillus. J. Ind. Microbiol. Biotechnol., 34: 241-245.
CrossRef - Ellaiah, P. and B. Srinivasulu, 1996. Production of extracellular protease by Streptomyces fradiae. Hindustan Antibiot. Bull., 38: 41-47.
PubMed - Felsenstein, J., 1981. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol., 17: 368-376.
CrossRefPubMedDirect Link - Hasegawa, T., M. Takizawa and S. Tanida, 1983. A rapid analysis for chemical grouping of aerobic actinomycetes. J. Gen. Applied Microbiol., 29: 319-322.
CrossRef - Hyo, J.K., C.L. Sung and K.H. Byung, 2006. Streptomyces cheonanensis sp. nov., a novel streptomycete with antifungal activity. Int. J. Syst. Evol. Microbiol., 56: 471-475.
Direct Link - Jensen, P.R., R. Dwight and W. Fenical, 1991. Distribution of actinomycetes in near-shore tropical marine sediments. Applied Environ. Microbiol., 57: 1102-1108.
Direct Link - Xu, J., Y. Wang, S.J. Xie, J. Xu, J. Xiao and J.S. Ruan, 2009. Streptomyces xiamenensis sp. nov., isolated from mangrove sediment. Int. J. Syst. Evol. Microbiol., 59: 472-476.
CrossRef - Keay, L. and B.S. Wildi, 1970. Proteases of genus Bacillus. I: Neutral proteases. Biotechnol. Bioeng., 12: 179-212.
PubMedDirect Link - Marmur, J. and P. Doty, 1962. Determination of the base composition of deoxyribonucleic acid from its thermal denaturation temperature. J. Mol. Biol., 5: 109-118.
CrossRefPubMedDirect Link - Minnikin, D.E., A.G. O'Donnell, M. Goodfellow, G. Alderson, M. Athalye, A. Schaal and J.H. Parlett, 1984. An integrated procedure for the extraction of bacterial isoprenoid quinones and polar lipids. J. Microbiol. Methods, 2: 233-241.
CrossRefDirect Link - El-Sersy, N.A., G.M. Abou-Elela, H. Abd-Elnaby, A.H.H. Ibrahim and N.M.K. El-Toukhy, 2010. Optimization, economization and characterization of cellulase produced by marine Streptomyces rubber. Afr. J. Biotechnol., 9: 6355-6364.
Direct Link - Shafee, N., S.N. Aris, R.N.Z.A. Rahman, M. Basri and A.B. Salleh, 2005. Optimization of environmental and nutritional conditions for the production of alkaline protease by a newly isolated bacterium Bacillus cereusm Strain 146. J. Applied Sci. Res., 1: 1-8.
Direct Link - Oskouie, S.F.G., F. Tabandeh, B. Yakhchali and F. Eftekhar, 2008. Response surface optimization of medium composition for alkaline protease production by Bacillus clausii. Biochem. Eng. J., 39: 37-42.
CrossRefDirect Link - Pridham, T.G. and D. Gottlieb, 1948. The utilization of carbon compounds by some Actinomycetales as an aid for species determination. J. Bacteriol., 56: 107-114.
Direct Link - Sen, S. and T. Satyanarayana, 1993. Optimization of alkaline protease production by thermophilic Bacillus licheniformis S-40. Indian J. Microbiol., 33: 43-47.
Direct Link - Seri, I.M., H. Haslinda, M. Mariatul Fadzillah, M.Y. Ishak and S.Z. Syed Osthman, 2005. Production of extra-cellular proteases from marine Bacillus sp. cultured in media containing ammonium sulfate as the sole nitrogen source. Malaysian J. Microbiol., 1: 30-34.
Direct Link - Qadar, S.A.U., E. Shireen, S. Iqbal and A. Anwar, 2009. Qptimization of protease production from newly isolated strain of Bacillus sp. PCSIR EA-3. Indian J. Biotechnol., 8: 286-290.
Direct Link - Shirling, E.B. and D. Gottlieb, 1966. Methods for characterization of Streptomyces species. Int. J. Syst. Evol. Microbiol., 16: 313-340.
CrossRefDirect Link - Staneck, J.L. and G.N. Roberts, 1974. Simplified approach to identification of aerobic actinomycetes by thin-layer chromatography. Applied Microbiol., 28: 226-231.
Direct Link - Sutcliffe, I.C., 2000. Characterisation of a lipomannan lipoglycan from the mycolic acid containing actinomycete Dietzia maris. Antonie van Leeuwenhoek, 78: 195-201.
CrossRefPubMedDirect Link - Li, W.J., Y.G. Zhang, Y.Q. Zhang, S.K. Tang, P. Xu, L.H. Xu and C.L. Jiang, 2005. Streptomyces sodiiphilus sp. nov., a novel alkaliphilic actinomycete. Int. J. Syst. Evol. Microbiol., 55: 1329-1333.
CrossRef - Williams, S.T., M. Goodfellow, G. Alderson, E.M.H. Wellington, P.H.A. Sneath and M.J. Sackin, 1983. Numerical classification of Streptomyces and related genera. J. Gen. Microbiol., 129: 1743-1813.
CrossRefPubMedDirect Link - Chen, X.G., O. Stabnikova, J.H. Tay, J.Y. Wang and S.T. Tay, 2004. Thermoactive extracellular proteases of Geobacillus caldoproteolyticus sp. nov., from sewage sludge. Extremophiles, 80: 489-498.
PubMed - Kuster, E. and S.T. Williams, 1964. Selection of media for isolation of Streptomycetes. Nature, 202: 928-929.
CrossRefDirect Link