
S. Omale
Department of Pharmacology and Toxicology, Faculty of Pharmaceutical Sciences University of Jos, Nigeria
LiveDNA: 234.33682
J.C. Aguiyi
Department of Pharmacology and Toxicology, Faculty of Pharmaceutical Sciences University of Jos, Nigeria
O.G. Adekunle
Department of Pharmacology and Toxicology, Faculty of Pharmaceutical Sciences University of Jos, Nigeria
T.O. Johnson
Department of Biochemistry, College of Medicine, University of Jos, Jos, Nigeria
S.O. Ochala
Drosophila Research Laboratory, Africa Centre of Excellence in Phytomedicine Research and Development, University of Jos, Jos, Nigeria
M.A. Etuh
Drosophila Research Laboratory, Africa Centre of Excellence in Phytomedicine Research and Development, University of Jos, Jos, Nigeria
M.C. Eze
Molecular Biology Research Laboratory, Africa Centre of Excellence in Phytomedicine Research and Development, University of Jos, Jos, Nigeria
Journal of Pharmacology and Toxicology
Year: 2021 | Volume: 16 | Issue: 1 | Page No.: 9-21







ABSTRACT
Background and Objective: Type 2 diabetes is a chronic metabolic disorder of insulin resistance or insulin insensitivity characterized by sustained hyperglycemia with microvascular and macrovascular complications. Adult individuals with diabetes have two to three-fold increased risks of heart attacks, stroke, neuropathy, foot ulcers and eventual limb amputation. Drosophila melanogaster (fruit-fly) has been used as a model for investigating human diseases because of its genetic resemblance to humans. This study investigated the antidiabetic and antioxidant effect of ethanolic extract of the stembark of Parinari curatellifolia in D. melanogaster (Harwich strain). Materials and Methods: High-calorie diet of sucrose (2.5 g/10 g diet) was given to Drosophila for ten days to induce insulin resistance type 2 diabetes. The diabetic flies were then treated with 0.5, 1.0, 2.0 and 4.0 mg of P. curatellifolia per 10 g diet for seven days, 16 mg/10 g diet of metformin was used as standard drug. The glucose content of the fly-homogenate was quantified using glucose oxidase. Results: Furthermore, we determined the total thiol contents, Nitric Oxide (NO) levels, as well as the activities of catalase (CAT), Acetylcholinesterase (AChE) and glutathione-S-transferase (GST). We observed a significant decrease (p<0.05) in the glucose content of diabetic flies treated with extracts and standard drugs. Conclusion: There was a substantial decrease in the total thiol contents, NO levels, AChE activity and a significant increase in GST and CAT activities of the flies. These data are consistent with antidiabetic agents’ effects, indicating P. curatellifolia as a potential remedy for insulin-resistant type 2 diabetes.
PDF Abstract XML References Citation
How to cite this article
S. Omale, J.C. Aguiyi, O.G. Adekunle, T.O. Johnson, S.O. Ochala, M.A. Etuh and M.C. Eze, 2021. Evaluation of the Antidiabetic Effects of the Stem Bark Extract of Parinari curatellifolia (Planch. ex Benth.) in Drosophila melanogaster. Journal of Pharmacology and Toxicology, 16: 9-21.
DOI: 10.3923/jpt.2021.9.21
URL: https://scialert.net/abstract/?doi=jpt.2021.9.21
DOI: 10.3923/jpt.2021.9.21
URL: https://scialert.net/abstract/?doi=jpt.2021.9.21
INTRODUCTION
Type 2 diabetes mellitus is a chronic metabolic disorder of insulin production or insensitivity characterized by sustained hyperglycemia with resultant long term microvascular (neuropathy, retinopathy and nephropathy) and macrovascular (coronary artery disease and cerebrovascular disease) complications1-3. There are three recognizable types of diabetes mellitus namely, Type I (formerly referred to as insulin-dependent), which is as a result of the failure of the pancreatic β-cells to produce insulin and is commonly found in children and adolescents, Type 2 (formally referred to as non-insulin-dependent) which is as a result of insulin resistance or insensitivity and Gestational diabetes which occurs during pregnancy. Type 2 diabetes is much more common and accounts for about 90% of cases of diabetes worldwide1,4.
An alarming increase in the prevalence of diabetes have been reported from 108 million in 1980-422 million in 2014 and the disease is the primary cause of blindness, heart attacks, kidney failure, stroke and lower limb amputation5. The International Diabetes Federation (IDF), estimated in 2015, 415 million adults will have diabetes and that, by 2040, the number would increase to 642 million6. In 2017, 451 million people (age 18-99) were living with diabetes worldwide, with a projected estimate of 693 million in 20457. Many countries have to spend 5-20% of national health budgets in 2015 to treat diabetes, with a global cost of 673 billion $. This global cost for diabetes treatment may rise to 802 billion $ by 20406 and 850 billion $ in 20457. Diabetes has been said to be the fifth leading course of death in the United States7. The incidence of diabetes is on the increase in Africa with a projected prevalence of 98% in 20 years and heavy burden on the national budgets of many countries8,9. Annual expenditure on diabetes in many countries of Africa has between put at 3.5-4.5 billion $10 and this figure is more than the total annual budgets of some African countries.
The search for alternative medicine to manage diabetes and other diseases has been on the African continent for decades. Several reports have shown that many medicinal plants are promising candidates in the management of diabetes. For example, bitter melon (Momordica charantia) has been shown to contain insulin-like substances that promote improved blood sugar control11. Extracts of E. jambolana and P. curatellifolia etc. have been shown to possess antidiabetic properties12-14.
World Health Organization (WHO)15 has continued to promote research into the use of substances from natural sources to manage chronic diseases such as diabetes since chemicals from organic sources are better tolerated by the human body16. As a result, copious researches are ongoing on the effectiveness and safety of medicinal products used in the maintenance of health in chronic diseases such as in high blood pressure17,18, Diabetes19-21 and Cancer22.
Among such plants with medicinal values is Parinari curatellifolia, Momordica charantia, Eugenia jambolana, some of which include blood glucose and low-density lipoprotein lowering activities12-14, anticancer23, anti-Snake venom24 and anti-hypertensive properties25. Traditionally it is also valued as food supplements; its sweet tasty fruits can be eating raw, cooked as porridge, syrups extracted as vegetable oil, or non-alcoholic drinks.
None of the medicinal properties of this plant has been investigated using fruit flies. In this study, we investigate for the first time in the Drosophila model the antidiabetic properties of this plant. Despite the wide application of D. melanogaster in biomedical research26-28, its use in ethnopharmacological screening of medicinal plants has not received the desired patronage. Drosophila melanogaster is a versatile, viable and affordable model that has been used in the study of various human diseases such as cancer29,30, cardiac diseases31, wound healing and inflammation32 and diabetes27,33.
Drosophila melanogaster was used as model for studying diabetes because it provides a useful complement to mammalian models and the receptors found in Drosophila have been shown to perform similar functions to insulin in mammals8,26,27,34. Drosophila insulin-like peptides have been shown to perform similar metabolic functions to mammalian insulin35,36. Drosophila has been recommended as an ideal model for metabolism studies and the mechanisms that maintain energy homeostasis, diabetes and obesity37.
This study aims to evaluate the antidiabetic and antioxidant effects of the stem bark extract of Parinari curatellifolia (SBEPC) on sucrose-induced Type 2 diabetes in D. melanogaster (fruit-fly).
MATERIALS AND METHODS
Study area: This study was carried out at the Drosophila Laboratory and the molecular biology Laboratory of Africa Centre of Excellence in Phytomedicine Research and Development (ACEPRD), University of Jos, Nigeria, from August, 2018-November, 2019.
Preparation of the plant material: The whole plant Parinari curatellifolia was collected in November, 2016 at about 11:00 am from its natural habitat in Oiji Ochekwu, Benue State, Nigeria and authenticated by a Taxonomist Mr. A. Owa, Head of Department, Agric-Extension Services and Federal College of Forestry Jos, Nigeria. An herbarium specimen was prepared and deposited in the Herbarium of Department of Pharmacognosy, Faculty of Pharmaceutical Sciences, University of Jos, Nigeria with the Voucher Specimen No. UJ/PCG/HSP/11C26.
The fresh stem bark of Parinari curatellifolia was air-dried at room temperature until sufficiently dried and pulverized into a coarse powder. Eight hundred grams (800 g) of the pulverized material was macerated in 70% ethanol for 72 hrs. The macerate was filtered using a Whatman filter paper (size 150 mm), Maidstone, England and the filtrate was concentrated in a water bath at 40°C.
Fly stock and culture: Wild type Drosophila melanogaster (Harwich strain) was obtained from the Drosophila research Laboratory of the Africa Centre of Excellence in Phytomedicine Research and Development (ACEPRD), University of Jos, Plateau State, Nigeria. The fly-stock was original from National Species Stock Centre (Bowling Green, OH, USA) but acquired by the Drosophila Laboratory, Department of Biochemistry, University of Ibadan, Nigeria, from which ACEPRD got its stock. All flies were maintained at a constant temperature of 25°C and 70% relative humidity with a 12 hrs light/dark clock cycle in vials containing standard cornmeal medium.
Young flies of 2-3 days old were collected under mild ice anesthesia from stock vials and divided into five groups of 50 flies (both gender). The flies were flipped into new vials containing fresh food every three days to ensure food quality consistency.
Survival assay: Effects of the extract on fly survival were studied; several pilot studies were carried out to arrive at the following conditions:
| • | Group I | : Control: 1 mL Ethanol/10 g diet |
| • | Group II | : About 0.5 mg of Parinari curatellifolia extract/10 g diet |
| • | Group III | : About 1.0 mg of Parinari curatellifolia extract/10 g diet |
| • | Group IV | : About 2.0 mg of Parinari curatellifolia extract/10 g diet |
| • | Group V | : About 4.0 mg of Parinari curatellifolia extract/10 g diet |
Concentrations of P. curatellifolia were made in the diet and fed to the flies. Each group consists of five replicates each and each replicate contained 50 flies. The flies were flipped to newly prepared diets containing the various concentrations every three days to maintain the diet’s consistency. Fly- mortality was recorded daily for twenty-eight days.
Induction of diabetes in flies: To induce type 2 diabetes in the flies, the method of Tennessen et al.37 was used with modifications. Briefly, sucrose (2.5 g sucrose/10 g diet) was incorporated into the regular fly diet to induce type 2 diabetes in wild type Harwich strain of Drosophila melanogaster. All other ingredients of the standard fly-food (1% agar, 3.4% yeast and, 8.3% cornmeal) were kept constant. Flies were observed after ten days for symptoms of diabetes, which include delayed egg production, delayed emergence of L3 Larvae, decreased body size for both larvae (L3) and adult flies and decreased locomotor activities. The glucose concentration of the fly- homogenate was quantified using the glucosidase method as described below.
Anti-diabetic activities of ethanolic stem-bark extract of Parinari curatellifolia in Drosophila melanogaster: To investigate the anti-diabetic effects of Parinari curatellifolia on Drosophila melanogaster, the methods described by Coogan35 were adopted with some modifications. Diabetic flies were placed on different concentrations of the stem bark extract and 16 mg metformin, respectively, for seven days, after which their glucose levels were determined. Seven groups of 50 flies (both gender) in 5 replicates were treated as follows:
| • | Group I | : Control (1 mL of ethanol) |
| • | Group II | : Diabetic flies untreated |
| • | Group III | : Diabetic flies treated with 16 mg metformin/10 g diet |
| • | Group IV | : Diabetic flies treated with 0.5 mg PC/10 g diet |
| • | Group V | : Diabetic flies treated with 1.0 mg PC/10 g diet |
| • | Group VI | : Diabetic flies treated with 2.0 mg PC/10 g diet |
| • | Group VII | : Diabetic flies treated with 4.0 mg PC/10 g diet |
Locomotor activities: The locomotor performance of both the diabetic and treated flies were determined using the negative geotaxis method as previously described38,39. Briefly, ten flies (both gender, five each) were immobilized under mild ice anesthesia placed separately in labeled vertical glass columns (length, 15 cm; diameter, 1.5 cm). After the recovery from ice exposure (about 20 min), the flies were gently tapped to the bottom of the column and the number of flies that climbed up to the 6 cm mark of the column in 6 seconds, as well as those that remained below the mark after this time, was recorded. Naturally, healthy flies move against gravity with preferring stay at the top of the glass column and occasionally move down to feed. The scores represent the mean of the number of flies at the top expressed as a percentage of the total number of flies. This procedure was repeated three times at 1 min interval and the mean score was used for the calculation.
Preparation of sample for biochemical assays: The samples for biochemical assays were prepared on day ten by anesthetizing the flies on ice. The flies were rinsed several times with cold potassium phosphate buffer solution (pH 7.4, 0.1 M) to remove all traces of food that might be attached to the flies37. Each group was deposited into pre-weighed Eppendorf tubes according to the various concentrations. The flies corresponding weight was recorded and the Phosphate Buffer Solution (PBS) at 10 μL buffer per mg fly body weight was added followed by homogenization in ice28,37. The homogenates were then centrifuged for 10 min at 4000 rpm (rotations per min.) in a Beckman Coulter, Inc Allegra X-I5R centrifuge, U.S.A that has been pre-chilled to 4°C. The resulting supernatants were pipetted into the corresponding pre-labeled Eppendorf tubes and stored at -18°C28. Subsequently, the supernatants were used to determine glucose, total protein, total thiol, nitric oxide levels, Acetylcholinesterase, Glutathione-S-Transferase, catalase activities.
Determination of glucose concentration: Randox Gluc-Pap assay kit, Randox Laboratory Crumlin County Antrim, United Kingdom, which uses the glucose oxidase method, was used in the determination of Glucose concentrations in the samples. The assay was carried out according to the manufacturer’s protocol.
The samples were diluted in 1:6 with PBS and mixed with the glucose oxidase reagents and incubated for 25 min at 25 and read at 540 nm using a Jenway 7315 spectrophotometer, Cole-Parmer Ltd., United Kingdom.
Total protein determination: The protein concentrations of the various samples were determined using the Bradford method40. This was carried out using the Randox protein assay kit, Randox Laboratory Crumlin County Antrim, United Kingdom, according to the manufacturer’s protocol. This is based on the binding of the protein molecules in the sample to Coomassie dye, which results in a color formation that can be read using a spectrophotometer. The reagents were mixed and incubated for 30 min at 25and the absorbance was measured at 546 nm using a spectrophotometer (Thermo Fisher Scientific). The standard curve was determined by plotting the absorbance against concentration.
The samples were mixed with the reagents and incubated for 30 min at 25 and the absorbance was measured at 546 nm using a spectrophotometer (Thermo Fisher Scientific).
Determination of the total thiol contents: The total thiols level was assayed according to the method of Ellman, as described by Abolaji et al.28 with some modifications. The total reaction mixture of 600 μL containing 25 μL of the sample, 510 μL potassium phosphate buffer (pH 7.4), 30 μL DTNB and 30 μL GSH. This was incubated for 30 min at 25°C and the absorbance was read at 412 nm using a spectrophotometer (Thermo Fisher Scientific). A GSH standard curve was prepared to extrapolate the total thiol content and the results were calculated per mg protein content.
Determination of acetylcholinesterase (AChE) activity: Acetylcholinesterase activity was assayed using the method of Ellman as described by Abolaji et al.28 with some modifications. The assay mixture was made up of 285 μL of distilled water, 180 μL of potassium phosphate buffer (pH 7.4), 60 μL of DTNB, 15 μL of the sample and 60 μL acetylthiocholine. The reaction was monitored for 2 min (15 sec intervals) at 412 nm using a spectrophotometer. The data were calculated against blank and sample blank and the results were corrected with the protein content.
Determination of catalase (CAT) activity: Catalase activity was determined according to the method of Claiborne41 with slight modifications. Briefly, 92 of 30% H2O2 was added to 25 mL of 0.1M phosphate buffer (pH 7.4) and the volume made up to 50 mL. The total reaction mixture was made up of 590 of H2O2 and 10 of the sample (1:60 dilutions PBS). The disappearance rate of H2O2 was read for 2 min (10 sec interval) at 240 nm using a spectrophotometer (Thermo Fisher Scientific).
Determination of nitric oxide (NO) level: Nitric oxide level was determined using Griess reagent as described by Bryan and Grisham42 with little modifications. Briefly, the Griess reagent was prepared using 50 mL of sulphanilamide solution, 50 mL of N-1-Naphthyl Ethylenediamine Dihydrochloride (NED) and 1 mL of 0.1 M Sodium nitrite. The reagents were mixed and stored at 4°C away from light until ready for use. Total 250 mL of sample in 250 mL of Griess reagent was incubated for 20 min and the absorbance was read at 550 nm.
Determination of glutathione-S transferase (GST) activity: The determination of the GST content of the sample was carried out according to the method described by Abolaji et al.28 with little modifications. Briefly, the assay reaction mixture was made up of 600 μL of a solution containing (20 μL mL of 0.25 M potassium phosphate buffer, pH 7.0, with 2.5 mM of ethylenediaminetetraacetic acid (EDTA), 10 μL of distilled water and 500 μL of 0.1 M GSH at 25°C, 60 μL of a sample (1:4 dilution) and 10 μL of 25 mM CDNB. The reaction was monitored for 2 min (10 s intervals) at 340 nm in a spectrophotometer.
Growth and developmental assay: To evaluate the effects of the high glucose diet on the mating, egg-laying, larva development and adult emergence and the rescue pattern of the extract of P. curatellifolia, we placed ten flies (same age) made up of five females and five males in seven groups as follows. Group, I Control (1 mL of ethanol), Group II 2.5 g Sucrose (diabetic flies), Group III Diabetic flies+16 mg metformin (standard), Group IV Diabetic flies+0.5 mg PC, Group V Diabetic flies+1 mg PC, Group VI Diabetic flies+2 mg PC, Group VII Diabetic flies+4 mg PC. All the groups were freshly prepared and allowed to mate and breed freely for 24 hrs in the breeding bottles during which they were observed four courtship and mating behaviors. The flies were transferred from the breeding bottles and the bottles containing eggs were observed for the appearance of first, second and third instar larva that eventually developed into a pupa and emerge as adult flies. The emergence of the flies was recorded daily for 14 days post-mating until the figures become constant for each group as no new flies continue to emerge.
Statistical analysis: The data were statistically analyzed using one-way ANOVA and multiple t-tests of Graph pad Prism version 7 and results are presented as mean±standard deviation. A 95% confidence interval was used to determine the statistical difference between the control and the treated and between different treated groups.
RESULTS
To evaluate the anti-diabetic effects of the extract of Parinari curatellifolia on the flies, we carried out several pilot studies to determine first the appropriate concentration of the extract to be used in the study. We found 0.5, 1, 2 and 4 mg/10 g diet to be safe for the treatment compared with the control. Secondly, we determined the concentration of the metformin to be used throughout the study to be 16 mg/10 g diet. To induce type 2 diabetes in the flies, we raised the flies on a fly diet containing high glucose content (2.5 g sucrose/10 g diet). The diabetic flies and larva (L3) were smaller in body size and showed decreased locomotor activities (Fig. 1).
Effects of varying concentrations of P. curatellifolia on fly survival: There was no statistical difference (p>0.05) in fly survival between the various concentrations of P. curatellifolia and the control ten days post-treatment. However, there was a gradual decrease in the fly beyond ten days when compared with the control. Conversely, there was a significant difference (p<0.05) between all the treated groups and the control at 28 days. The extracts at 0.5, 1, 2 and 4 mg/10 g diet decreased the survival rate of flies by 80.4, 83.6, 80.8 and 81.2% respectively compared with 91.6% survival rate in the control. There was no significant difference (p>0.05) in the survival rate of the flies between the various concentrations (Fig. 2).
Locomotor activities: The locomotor performance of the flies was assayed using the climbing method. Diabetic flies were found to be sluggish with reduced activities, such as the inability to climb up towards light (negative geotaxis). The diabetic fly locomotor performance treated with 0.5, 1 and 2 mg was significantly improved (p<0.05) compared with the control. The highest locomotor performance of the flies was observed in the groups treated with 1 mg of the extract of P. curatellifolia (Fig. 3).
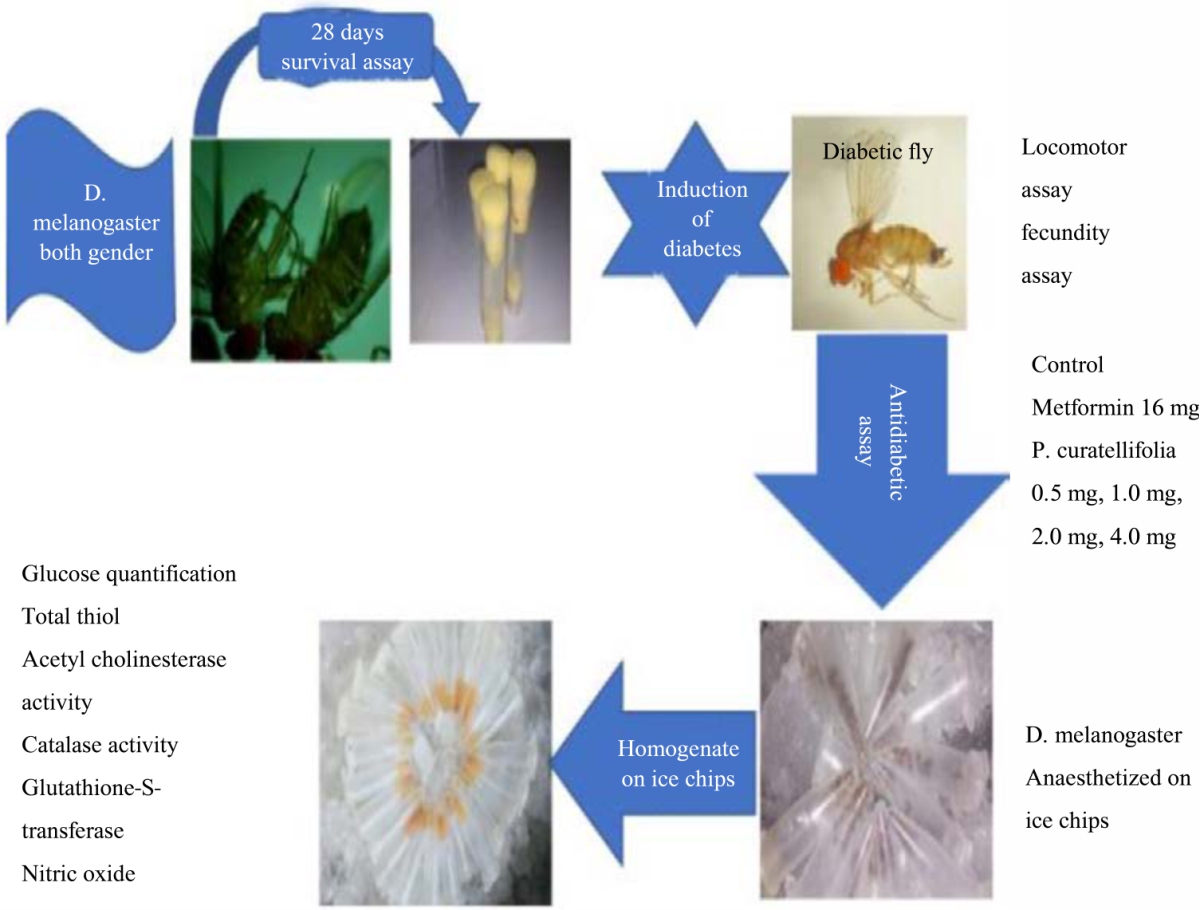 |
| Fig. 1: | Scheme of experimental design showing research flow chart |
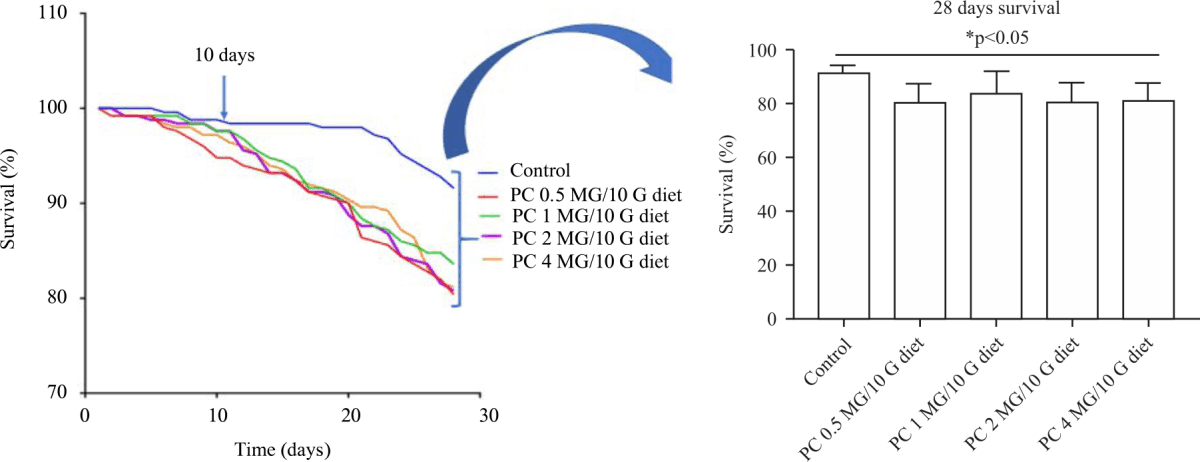 |
| Fig. 2: | Life span of Drosophila melanogaster starts decreasing shortly after 10 days of treatment with 0.5, 1, 2 and 4 mg/10 g diet There was a significant decrease (p<0.05) in the life span of the flies beyond 10 days compared with the control. However, there was no significant difference (p>0.05) in the life span of the flies within treatment groups. Overall, the effects of the different concentrations of the extract are within safety margin as reflected by 80.4% (0.5 mg), 83.6% (1 mg), 80.8% (2 mg) and 81.2% (4 mg) fly survival after 28 days treatment |
Hypoglycemic effect of the stem-bark extract of Parinari curatellifolia on type 2 diabetes in Drosophila melanogaster: The effects of the various concentrations of the ethanolic extract of P. curatellifolia on the glucose levels were assayed after ten days of high sucrose diet. The flies were observed to have decreased body size, decreased locomotor activities, delayed emergence of the larva (L3), which are some of the symptoms of type 2 diabetes in the flies. We assayed anti-diabetic effects of the four different concentrations (0.5, 1, 2 and 4 mg) of the extract of P. curatellifolia and the effects of metformin (16 mg), an oral hypoglycemic drug on the diabetic flies.
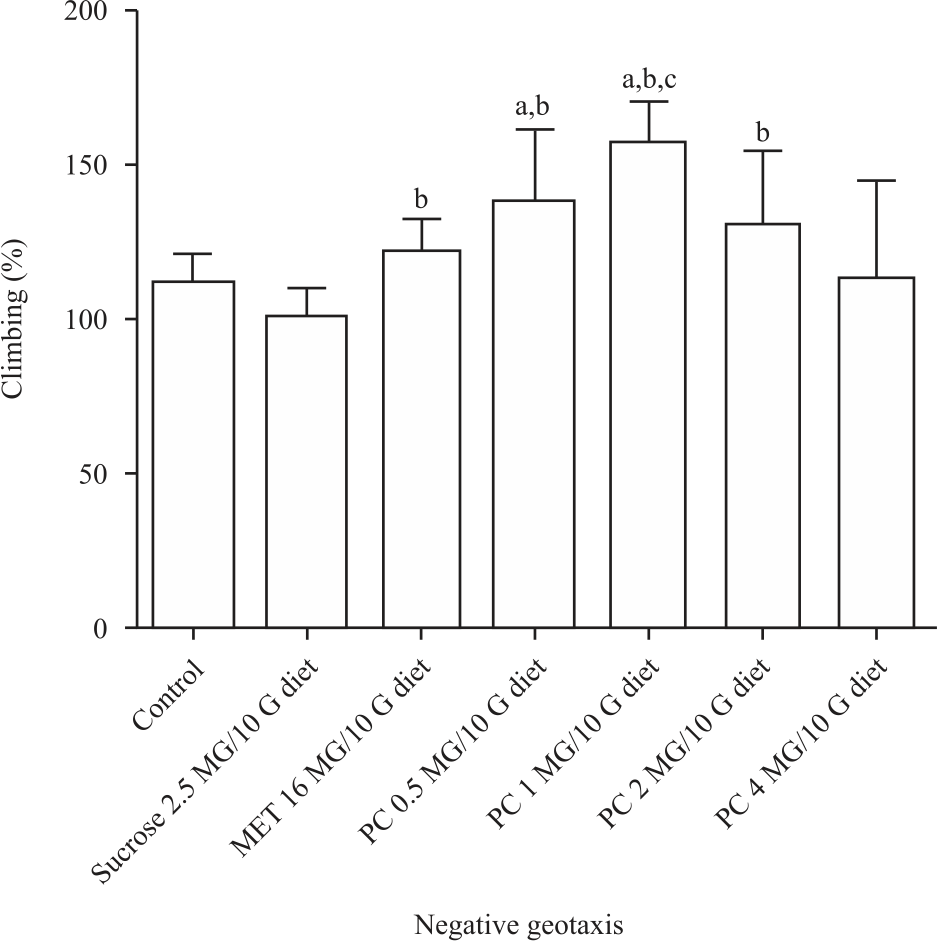 |
| Fig. 3: | Extract of P. curatellifolia significantly increased (p<0.05) the locomotor activities of the diabetic flies treated with 0.5, 1 and 2 mg/10 g diet a: Statistical difference between the treatment group and the control group, b: Statistical different between the treatment group and the high sucrose group, c: Statistical different within the treated groups. There was no proportionate increase in locomotor activities of the flies treated with 4 mg/10 g diet. This observation could be due to a higher concentration of the extract tending towards overdose as the toxicity of every drug is dependent on the concentration |
The experiment consist of seven groups of 50 flies in 5 replicate each; Group 1 (regular fly diet), group 2 (2.5 g sucrose only), group 3 (2.5 g sucrose+metformin 16 mg) and groups 4, 5, 6 and 7 (2.5 g sucrose+0.5 mg, 2.5 g sucrose+1 mg, 2.5 g sucrose+2 mg and 2.5 g sucrose+4 mg extract of P. curatellifolia, respectively). We used the glucosidase method to quantify the glucose levels of each group. The results showed normal glucose content in group 1 (normal fly diet) and high glucose level (p<0.05) in group 2 compared with the control. The extracts of P. curatellifolia at the tested concentrations significantly decreased (p<0.05) the glucose levels of groups 4, 5, 6 and 7 with the greatest decrease in group 6 (2 mg of the extract) compared with the diabetic control (group 3). All different concentrations of the extract reduce glucose level, similar to metformin 16 mg (Fig. 4).
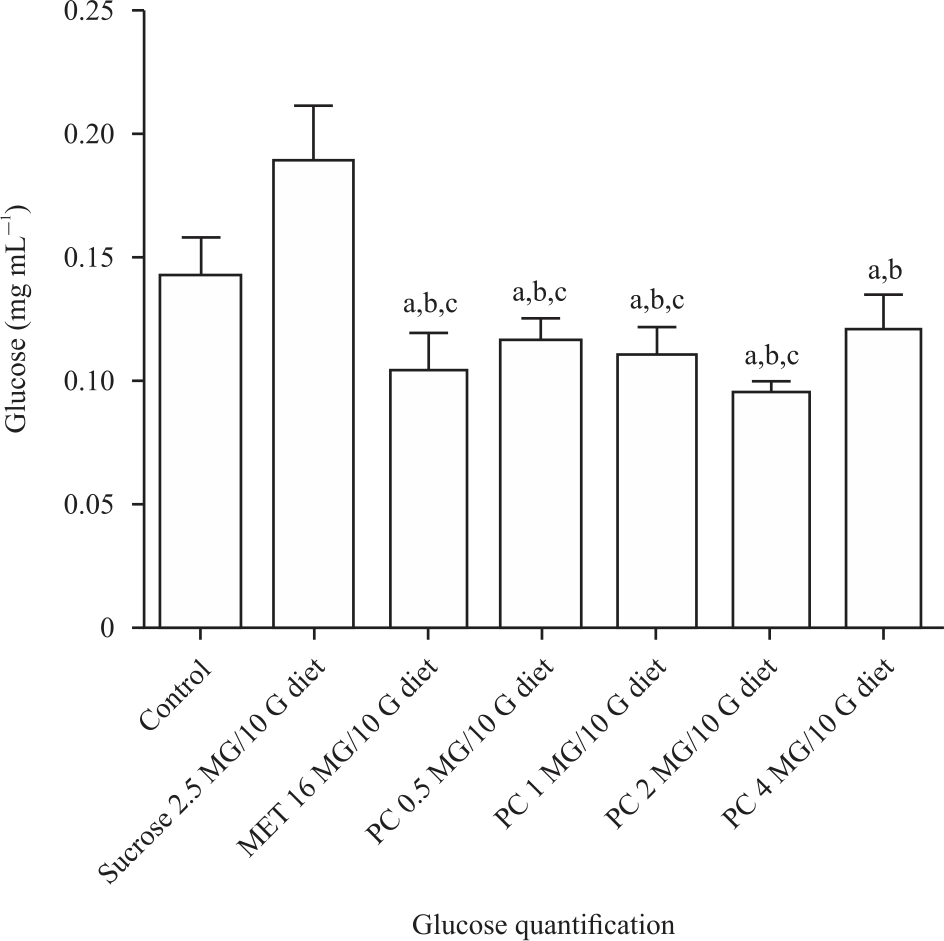 |
| Fig. 4: | Glucose level of type 2 diabetic Drosophila melanogaster treated with 0. 5, 1, 2 and 4 mg of the extract of P. curatellifolia and metformin 16 mg after 10 days a: Statistical difference between the treatment group and the control group, b: Statistical different between the treatment group and the high sucrose group, c: Statistical different within the treated groups. All groups contain 50 flies each in 5 replicates and data are presented as Mean±SD. There was a significant decrease in the glucose content of all the treated groups compared with the diabetic control |
Stem-bark extract of P. curatellifolia decreased total thiol, NO and AChE but increased GSH and CAT activity: The extract of P. curatellifolia significantly decreased (p<0.05) the total thiol content (an oxidative stress marker) of the diabetic flies when compared with the control (Fig. 5a). The nitric oxide level of the diabetic D. melanogaster was also reduced in the groups treated with 0.5, 2 and 4 mg of the extract, respectively (Fig. 5b). There was a significant decrease (p<0.05) in the acetylcholinesterase activities of all the groups treated with the extract of P. curatellifolia (Fig. 5c). There was a significant increase (p<0.05) in the glutathione-S-transferase and catalase activity in the diabetic D. melanogaster treated with 0.5, 1, 2 and 4 mg of the extract of P. curatellifolia (Fig. 5d-e).
Extract of P. curatellifolia increased mating, egg-laying, larva development and fly emergence: We observed an increased courtship and mating behavior in the flies treated with the extract of P. curatellifolia compared with diabetic control.
 |
| Fig. 5: | Extracts of P. curatellifolia significantly reduced (p<0.05) the total thiol content of sucrose induced type 2 diabetes in wild type D. melanogaster (a) A high level of total thiols in the hyperglycemic flies is an indication of oxidative stress in type 2 diabetes which was rescued in the groups treated with extracts of P. curatellifolia, (b) Nitric oxide (NO) is a free radical reactive oxygen species, that when in high concentration during inflammation can cause tissue damage. However, it is a potent vasodilator that is usually impaired in type 2 diabetes. We observed a significant decrease (p<0.05) in nitric oxide activities of all the treated groups compared with the control, (c) A decreased AChE activity was observed in all the groups compared with the control, (d) Extract of P. curatellifolia significantly (p<0.05) increased the glutathione-S-transferase (GST, an endogenous antioxidant enzyme) activity in the type 2 diabetic D. melanogaster and (e) Extract of P. curatellifolia (0.5, 1, 2 and 4 mg) significantly increased (p<0.05) catalase (CAT, an endogenous antioxidant enzyme) activity in the type 2 diabetic D. melanogaster. a: Statistical difference between the treatment group and the control group, b: Statistical different between the treatment group and the high sucrose group, c: Statistical different within the treated groups |
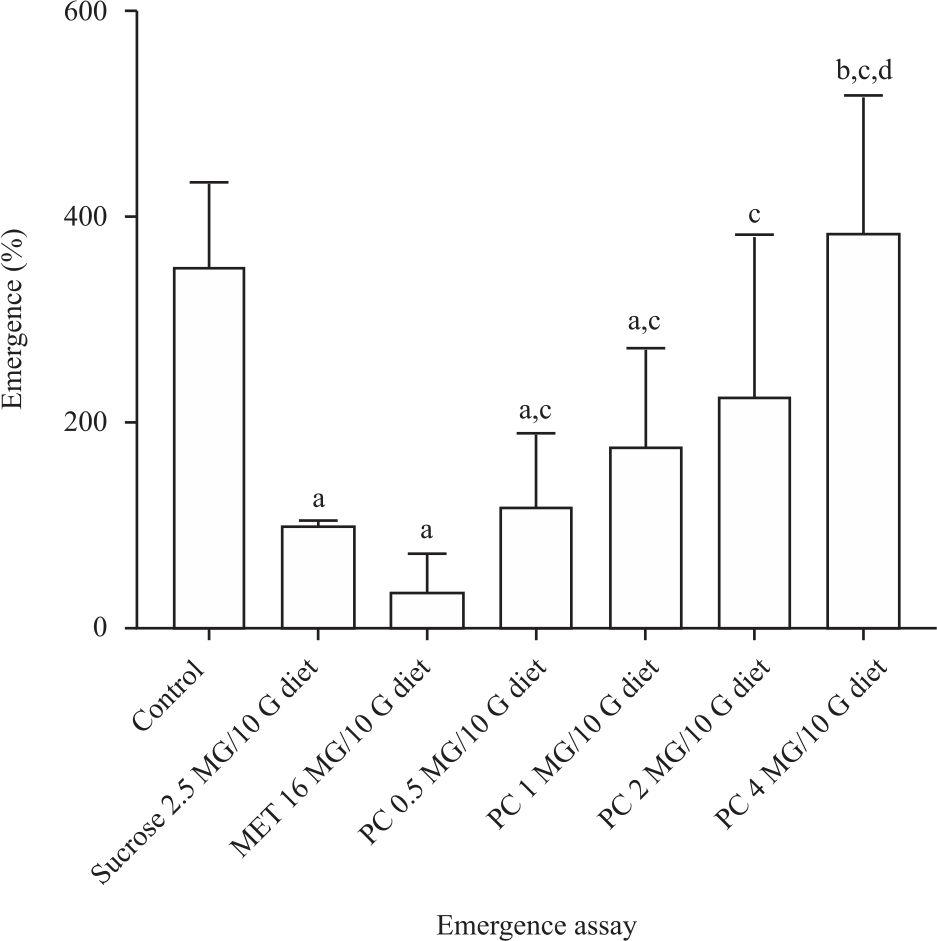 |
| Fig. 6: | Extract of P. curatellifolia significantly increased (p<0.05) egg-laying, mating behavior, larva development and adult emergence in treated groups of sucrose induced type 2 diabetic D. melanogaster compared with the untreated a: Statistical difference between the treatment group and the control group, b: Statistical different between the treatment group and the high sucrose group, c: Statistical different within the treated groups |
The life cycle from egg to adult flies of the diabetic group was longer than the treated group, as signifies by the early emergence of L3 and adult flies in the treated groups (data not presented). There was a dose-dependent increase (p<0.05) in the number of adult flies that emerge 14 days post-mating (Fig. 6).
DISCUSSION
The effects of 0.5, 1, 2 and 4 mg of the extract of P. curatellifolia on D. melanogaster’s lifespan showed an adequate level of safety as all the concentrations scored over 80% survival at the end of 28 days of treatment (Fig. 2). There is no significant difference (p>0.05) between the control and the treated groups. As obtained from this experiment, the extract's safety profile showed that the extract is safe, as reported by the previous researchers13,14,24,25.
The locomotor activities showed a significant decrease (p<0.05) in locomotor performance in the diabetic group compared with the control. However, an improved locomotor activity (p>0.05) was observed in the treated groups compared with the control (Fig. 3). It was also observed that the increased muscular activities were higher at 0.5, 1 and 2 mg doses of the extract than the metformin control. This implies that the activity of the extract at this point could be higher than metformin. Insulin resistance diabetes has been reported to have peripheral neuropathy symptoms such as increased muscle weakness, loss of reflexes especially of the legs and loss of balance and coordination43. The decreased locomotor activities seen in the diabetic group could be attributed to these symptoms of diabetic neuropathy. For the first time, we investigated the ability of the extract of P. curatellifolia to reverse the neuromuscular deficits of type 2 diabetes in D. melanogaster.
The feeding of the flies with high sucrose (2.5 g/10 g diet) resulted in insulin-resistant diabetes typified by decrease in size of both adult flies and larvae8 compared with the control group. Both adult flies and the wandering L3 larvae were also observed to have very sluggish movements, which were attributed to a neuromuscular defect of type 2 diabetes. We quantify the flies’ glucose content using the glucosidase method and found a high glucose content in the flies fed with high sucrose diet alone compared with the control. In the treatment groups with 0.5, 1, 2 and 4 mg of P. curatellifolia and 16 mg metformin, a corresponding decrease (p<0.05) in glucose content was observed. Metformin (a biguanide) is an oral hypoglycemic agent and insulin sensitizer is a common choice in the management of insulin-resistant diabetes; this is why we choose it in this experiment. We observed that there was no significant difference (p>0.05) in the extent of the reduction in glucose level between metformin and the extract of P. curatellifolia (Fig. 4). From these observations, we suggest that metformin’s mode of action and, the extract of P. curatellifolia could be similar. We also observed a similar pattern in the rescued body size and locomotor activities for both the larvae and adult flies in the metformin and the groups treated with the extract. For the first time, we have shown that the extract of P. curatellifolia has hypoglycemic effects on insulin resistance diabetes in D. melanogaster.
In this present study, we investigate insulin-resistant diabetes’ effects on some oxidative stress biomarkers (such as total thiols, glutathione-S-transferase, catalase, nitric oxide and acetylcholinesterase) and the antioxidant effects of the extract of P. curatellifolia in Drosophila. We observed high levels of total thiols in the diabetic group consistent with oxidative stress in Drosophila; conversely, a decreased level (p<0.05) of total thiol was observed in the 0.5, 1 and 4 mg and the metformin control (Fig. 5a). Indeed, oxidative stress has been implicated in the development and complications of many chronic human pathologies such as diabetes, obesity, cancer, cardiovascular diseases, Alzheimer's disease and aging44,45.
Oxidation of animal cells ‘components can result in cellular damage leading to various pathologies; antioxidants response to oxidation is a physiological defensive mechanism8. There is an increased level of antioxidant biomarkers such as total thiol, glutathione-S-transferase, superoxide dismutase, hydrogen peroxide and antioxidant enzymes such as catalase, Acetylcholinesterase during oxidative stress46.
Metformin has been demonstrated to have decreased effects on oxidative stress markers such as total thiol, catalase, glutathione-S-transferase47,48. Based on these results, we speculate that metformin and extract of P. curatellifolia possess similar antioxidant activity by reducing the total thiol level in insulin resistance diabetic D. melanogaster.
We also observed a decreased Nitric oxide and Acetylcholinesterase level in all the treated groups compared with the control (Fig. 5b-c). This is consistent with other findings that nitric oxide is inactivated in type 2 diabetic patients49. Acetylcholinesterase (AChE) terminates the action of the physiological Acetylcholine neurotransmitter. The decreased AChE activity observed in this experiment is thought to be the body’s adaptive strategy in sparing the available Acetylcholine to preserve the neuromuscular activities of the fly (Fig. 5c).
In this study, we observed an increased level of GST activity in the diabetic group as a natural body response, which is augmented in the treated groups with 0.5, 2 and 4 mg of the extract (Fig. 5d). We attribute the high levels of GST observed in the treated groups to be increased antioxidant effects of the extract of P. curatellifolia to support the body’s defense against oxidative stress. A positive correlation exists between glutathione-S-transferase activity and the complications of Type 2 diabetes, such as diabetic neuropathy50. Glutathione-S-transferase is a potent endogenous antioxidant enzyme that conjugates reactive oxygen species and neutralizes the harmful effects of oxidative stress. In type 2 diabetes, hyperglycemia induces oxidative stress as a result of increased Reactive Oxygen Species (ROS) and Reactive Nitrogen Species (RNS). The body’s natural defensive response to increased ROS and RNS is increased production of antioxidants51.
The results showed increased catalase activity (p<0.05) in the groups treated with metformin and the extracts of P. curatellifolia compared with diabetic untreated groups, while there was no significant difference (p>0.05) between the metformin and the extract-treated groups (Fig. 5e). Deficiency of catalase (CAT) an endogenous antioxidant enzyme involved in the conversion of hydrogen peroxide (H2O2) to oxygen and water, has been observed in type 2 diabetes52. The observation in our metformin and treated groups are consistent with published data that metformin improves catalase activity in mice challenged with lipopolysaccharide/D-galactosamine (LPS/D-Gal)-induced endotoxic liver injury53. The extract of P. curatellifolia significantly increased (p<0.05) catalase activities in type 2 diabetes in Drosophila melanogaster and thus acts as potent antioxidant phytochemicals.
We demonstrate the ability of the extract of P. curatellifolia to rescue the effects of type 2 diabetes on mating behavior, egg-laying, wandering L3 larva and adult emergence. The wandering third instar larvae in the diabetic group were observed to appear late (6-7 days) than the control (5-6) days. We also marked the late emergence of the adult flies in the diabetic group. In contrast, in the treated groups, the third instar larvae appeared early (5-6) days and have a normal size with improved locomotor activities. There was a dose-dependent increase (p<0.05) in the number of adult flies that emerge in the treatment groups at the end of 14 days (Fig. 6). The number of adult flies that emerge at the end of the treatment was about threefold compared with the diabetic group. Type 2 diabetes has been shown to affects all stages of the Drosophila life cycle with adults mating less and fewer egg-laying with delayed appearance of third instar larvae in the life cycle8. We conclude that the extracts of P. curatellifolia dramatically improves growth and development in type 2 diabetic flies at all the tested concentrations.
This study’s implication is the justification of the use of the stem bark extract of P. curatellifolia by alternative medicine practitioners. This study also showed the extract’s safety and the ability to reverse some complications of Type 2 diabetes, such as neuromuscular and fecundity deficits. This study’s application is the scientific evidence of the use of P. curatellifolia by traditional medicine practitioners in the management of Type 2 diabetes.
The limitation of this study is the inability to establish the specific mechanism of the extract’s antidiabetic activities. The study was not able to characterize the active principles responsible for the antidiabetic effects. Therefore, we recommend further evaluation and characterization of the medicinal plant’s active principles as possible raw material for pharmaceutical industries.
CONCLUSION
From our observations, we conclude that the ethanolic extract of the stembark of Parinari curatellifolia rescued the neuromuscular defects and restored normal glucose levels in type 2 diabetes in Drosophila melanogaster. We also conclude that type 2 diabetes induces oxidative stress in D. melanogaster and that the extracts of P. curatellifolia have remarkable antioxidant effects in the flies. Furthermore, the extracts of P. curatellifolia dramatically improved growth and development in type 2 diabetic flies. Overall, the extract of P. curatellifolia has a demonstrable antioxidant and anti-diabetic effects on type 2 diabetes in D. melanogaster (fruit-fly).
SIGNIFICANCE STATEMENT
This study substantiated the antidiabetic effects of the extract and discovered the specific activities of the antioxidant markers involved in the antidiabetic effects of the stem bark extract of P. curatellifolia using Drosophila. This can be beneficial in understanding the mechanism of action of the plant’s constituents. This study will help the researcher develop antidiabetic and other drugs from plant origin using the Drosophila melanogaster model, which many researchers of ethnopharmacology were not as now able to explore.
ACKNOWLEDGMENTS
The authors acknowledged Mr. Omale Ikwuoche of Oiji ochekwu, Benue State, Nigeria, who provided the ethnomedicinal knowledge of the plant. The authors are also grateful to all the staff of the Drosophila laboratory of Africa Centre of Excellence in Phytomedicine Research and Development (ACEPRD) for the use of their laboratory and equipment for this research.
REFERENCES
- Ogbera, A.O., 2014. Diabetes mellitus in Nigeria: The past, present and future. World J. Diabetes, Vol. 5.
CrossRefDirect Link - Gupta, H.P., R.R. Jha, H. Ahmad, D.K. Patel and K.R. Ram, 2019. Xenobiotic mediated diabetogenesis: Developmental exposure to dichlorvos or atrazine leads to type 1 or type 2 diabetes in Drosophila. Free Radical Biol. Med., 141: 461-474.
CrossRefDirect Link - Micheal, J. and M.D. Fowler, 2011. Microvascular and macrovascular complications of diabetes mellitus. Clin. Diabetes, 29: 116-122.
CrossRefDirect Link - Zheng, Y., S.H. Ley and F.B. Hu, 2018. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol., 14: 88-98.
CrossRefDirect Link - Standl, E., K. Khunti, T.B. Hansen and O. Schnell, 2019. The global epidemics of diabetes in the 21st century: Current situation and perspectives. Eur. J. Prev. Cardiol., 26: 7 -14.
CrossRefDirect Link - Ogurtsova, K., J.D. da Rocha Fernandes, Y. Huang, U. Linnenkamp and L. Guariguata et al., 2017. IDF Diabetes Atlas: Global estimates for the prevalence of diabetes for 2015 and 2040. Diabetes Res. Clin. Pract., 128: 40-50.
CrossRefDirect Link - Cho, N.H., J.E. Shaw, S. Karuranga, Y. Huang, J.D. da Rocha Fernandes, A.W. Ohlrogge and B. Malanda, 2018. IDF diabetes atlas: Global estimates of diabetes prevalence for 2017 and projections for 2045. Diabetes Res. Clin. Pract., 138: 271-281.
CrossRefDirect Link - Musselman, L.P., J.L. Fink, K. Narzinski, P.V. Ramachandran, S.S. Hathiramani, R.L. Cagan and T.J. Baranski, 2011. A high-sugar diet produces obesity and insulin resistance in wild-type Drosophila. Dis. Model. Mech., 4 : 842-849.
CrossRef - Jaffiol, C., 2011. The burden of diabetes in Africa: A major health problem. Bull. Acad. Nat. Med., 196: 1239-1253.
CrossRefDirect Link - Mutyambizi, C., M. Pavlova, L. Chola, C. Hongoro and W. Groot, 2018. Cost of diabetic mellitus in Africa: A systemic review of existing literature. J. Glob. Health, 14 : 1186-1198.
CrossRefDirect Link - Srivastava, Y., H. Venkatakrishna-Bhatt, Y. Verma, K. Venkaiah and B.H. Raval, 1993. Antidiabetic and adaptogenic properties of Momordica charantia extract: An experimental and clinical evaluation. Phytother. Res., 7: 285-289.
Direct Link - Ravi, K., B. Ramachandran and S. Subramanian, 2004. Protective effect of Eugenia jambolana seed kernel on tissue antioxidants in streptozotocin-induced diabetic rats. Biol. Pharm. Bull., 27: 1212-1217.
CrossRefDirect Link - Ogbonnia, S.O., A.A. Adekunle, S.O. Olagbende-Dada, E.N. Anyika, V.N. Enwuru and M. Orolepe, 2008. Assessing plasma glucose and lipid levels, body weight and acute toxicity following oral administration of an aqueous ethanolic extract of Parinari curatellifolia Planch (Chrysobalanceae) seeds in alloxan-induced diabetes in rats. Afr. J. Biotech., 7: 2998-3003.
Direct Link - Ogunbolude Y., M.A. Ajayi, T.M. Ajagbawa, A.P. Igbakin, J.B.T. Rocha and I.J. Kade, 2009. Ethanolic extracts of seeds of Parinari curatellifolia exhibit potent antioxidant properties: A possible mechanism of its antidiabetic action. J. Pharmacog. Phytoth., 1: 67-75.
Direct Link - Mahomoodally, M.F., 2013. Traditional medicines in Africa: An appraisal of ten potent African medicinal plants. Evidence-Based Complementary Altern. Med., Vol. 2013.
CrossRefDirect Link - Tilburt, J.C. and T.J. Kaptchuk, 2008. Herbal medicine research and global health: An ethical analysis. Bull. World Health Organ., 86: 594-599.
Direct Link - Tahraoui, A., J. El-Hilaly, Z.H. Israili and B. Lyoussi, 2007. Ethnopharmacological survey of plants used in the traditional treatment of hypertension and diabetes in South-Eastern Morocco (Errachidia province). J. Ethnopharmacol., 110: 105-117.
CrossRefPubMedDirect Link - Yuliandra, Y., A. Armenia and H. Arifin, 2017. Antihypertensive and anti-oxidant activity of Cassytha filiformis L: A correlative study. Asian Pac. J. Trop. Biomed., 7: 614-618.
CrossRefDirect Link - Husain, G.M., P.N. Singh and V. Kumar, 2009. Antidiabetic activity of standardized extract of Picrorhiza kurroa in rat model of NIDDM. Drug Discoveries Therapeutics, 3: 88-92.
PubMedDirect Link - Bhagwat, D.A., S.G. Killedar and R.S. Adnaik, 2008. Anti-diabetic activity of leaf extract of Tridax procumbens. Int. J. Green Pharm., 2: 126-128.
Direct Link - Yimam, M., J. Zhao, B. Corneliusen, M. Pantier, L. Brownell and Q. Jia, 2014. Blood glucose lowering activity of aloe based composition, UP780 in alloxan induced insulin-dependent mouse diabetic model. Diabetol. Metab. Syndr., Vol. 6.
CrossRefDirect Link - Al-Asmari, A.K., S.M. Albalawi, M.T. Athar, A.Q. Khan, H. Al-Sharani and M. Islam, 2015. Moringa oleifera as an anti-cancer agent against breast and colorectal cancer cell lines. PLoS One, Vol. 10.
CrossRefDirect Link - Rundle, N.T., X. Lin, J.A. Raymond and R. Micheal, 2011. G2 DNA damage checkpoint inhibition and antimitotic activity of 13- hydroxy-15-oxozoapatin from Parinari curatellifolia. J. Biol. Chem., 276: 48231-48236.
CrossRefDirect Link - Omale, S., A. Asa, K.I. Amagon and V.I. Mariose, 2012. Anti-snake venom activity of flavonoids from the root bark extract of Parinari curatellifolia in mice. Int. J. Pharm. Res., 4 : 55-58.
Direct Link - Omale, S., A. Auta, S.B. Banwat, K.I. Amagon and Y.P. Thomas, 2011. Effects of the ethanolic extract of Parinari curatellifolia on cat blood pressure and rabbit jejunum preparations. Int. J. Pharm. Frontier Res., 1: 39-44.
Direct Link - Morris, S.N.S., C. Coogan, K. Chamseddin, S.O. Fernandez-Kim, S. Kolli, J.N. Keller and J.H. Bauer, 2012. Development of diet-induced insulin resistance in adult Drosophila melanogaster. Biochim Biophys. Acta, 1822: 1230-1237.
CrossRefDirect Link - Graham, P. and L. Pick, 2017. Drosophila as a model for diabetes and diseases of insulin resistance. Curr. Top. Dev. Biol., 121: 397-419.
CrossRefDirect Link - Abolaji, A.O., C.O. Olaiya, O.J. Oluwadahunsi and E.O. Farombi, 2017. Dietary consumption of monosodium L-glutamate induces adaptive response and reduction in the life span of Drosophila melanogaster. Cell Biochem. Funct., 35: 164-170.
CrossRefDirect Link - Halder, G. and G.B. Mills, 2011. Drosophila in cancer research: To boldly go where no one has gone before. Oncogene, 30: 4063-4066.
CrossRefDirect Link - Vivek, A.R., L.C. Ross and K.D. Tirtha, 2012. Drosophila cancer models. Dev. Dyn., 241: 107-118.
CrossRefDirect Link - Piazza, N. and R.J. Wessells, 2011. Drosophila models of cardiac diseases. Prog. Mol. Biol. Transl. Sci., 100: 155-210.
CrossRefDirect Link - William, R., W. Will and M. Paul, 2011. Swatting flies: Modelling wound healing and inflammation in Drosophila. Dis. Model. Mech., 4 : 569-574.
CrossRefDirect Link - Read, R.D., P.J. Goodfellow, N.N. Mardis, J.R. Armstrong and R.L. Cagan, 2005. A Drosophila model of multiple endocrine neoplasia type II. Genetics, 171: 1057-1081.
CrossRefDirect Link - Alfa, R.W. and S.K. Kim, 2016. Using Drosophila to discover mechanisms underlying type 2 diabetes. Dis. Model. Mech., 9: 365-376.
CrossRefDirect Link - Coogan, C.M., 2013. Diagnosis and prevention of metabolic diseases in Drosophila melanogaster. Engaged Learn. Collect., Vol. 27.
Direct Link - Pasco, M.Y. and P. Leopold, 2012. High sugar-induced insulin resistance in Drosophila relies on the lipocalin neural lazarillo. PLoS ONE, Vol. 7.
CrossRefDirect Link - Tennessen, J.M., W. Barry, J. Cox and C.S. Thummel, 2014. Methods for studying metabolism in Drosophila. Methods, 68: 105-115.
CrossRefDirect Link - Feany, M.B. and W.W. Bender, 2000. A drosophila model of Parkinson's disease. Nature, 404: 394-398.
PubMed - Adedara, I.A., A.O. Abolaji, J.B.T. Rocha and E.O. Farombi, 2016. Diphenyl diselenide protects against mortality, locomotor deficits and oxidative stress in Drosophila melanogaster model manganese-induced neurotoxicity. Neurochem. Res., 41: 1430-1438.
CrossRefDirect Link - Kruger, N.J., 1996. The Bradford Method for Protein Quantitation. In: The Protein Protocols Handbook, Walker, J.M. (Ed.). Humana Press, New Jersey, pp: 15-20.
CrossRefDirect Link - Greenwald, R.A., 1985. CRC Handbook of Methods for Oxygen Radical Research. 1st Ed. CRC Press, Boca Raton, ISBN-13: 9780849329364, pp: 267-278.
Direct Link - Bryan, N.S. and M.B. Grisham, 2007. Methods to detect nitric oxide and its metabolites in biological samples. Free Radical Biol. Med., 43: 645-657.
CrossRefDirect Link - Bansal, V., J. Kalita and U.K. Misra, 2006. Diabetic neuropathy. Postgrad. Med. J., 82: 95-100.
CrossRefDirect Link - Giacco, F., M. Brownlee and A.M. Schmidt, 2010. Oxidative stress and diabetic complications. Circ. Res., 107: 1058-1070.
CrossRefPubMedDirect Link - Matschke, V., C. Theiss and J. Matschke, 2019. Oxidative stress: The lowest common denominator of multiple diseases. Neural Regen. Res., 14: 238-241.
CrossRef - Ravikurma, H.M., 2013. Acute exposure of Drosophila melanogaster to paraquat causes oxidative stress and mitochondrial dysfunction. Arch. Insect Biochem. Physiol., 83: 25-40.
CrossRefDirect Link - Alireza, E., E. Delaram, M. Hossein, N. Sina, M. Mostafa, H. Mehdi and N. Manoucherh, 2013. Effects of metformin on markers of oxidative stress and antioxidant reserve in patients with newly diagnosed type 2 diabetes: A randomize clinical trial. Clin. Nutr., 32: 179-185.
CrossRefDirect Link - Obi, B.C., T.C. Okoye, V.E. Okpashi, C.N. Igwe and E.O. Alumanah, 2016. Comparative study of the antioxidant effects of metformin, glibenclamide and repaglinide in alloxan-induced diabetic rats. J. Diabetes Res. Vol. 2016.
CrossRefDirect Link - Huszka, M., M. Kaplar, L. Rejto, I. Tornai, K. Palatka, P. Laszlo and M. Udvardy, 1997. The association of reduced endothelium derived relaxing factor-no production with endothelial damage and increased in vivo platelet activation in patients with diabetes mellitus. Thromb. Res., 86: 173-180.
CrossRefDirect Link - Mohini, S., G. Stuti, S. Kalpana, M. Mohit and G. Amar, 2016. Association of glutathione -s-transferase with patient of type 2 diabetes mellitus with and without neuropathy. Diabetes Metab. Syndr., 10: 194-197.
CrossRefDirect Link - Laura, R.B.A, C.S.R. Ana, R.P. Gustavo, S.S. Rodrigo and A.S.R. Angela, 2018. Impact of oxidative changes and possible effects of genetics polymorphisms of glutathione S-Transferase in diabetics patients with complications. In: Glutathione in Health and Disease, Erkekoglu, P. and B. Kocer-Gumusel, BoD-Books on Demand, IntechOpen, ISBN: 9781789842753 Pages: 92.
CrossRefDirect Link - Goth, L., 2008. Catalase deficiency and type 2 diabetes. Diabetes Care, 31: e93-e93.
CrossRefPubMedDirect Link - Ai, Q., T. Dai. M.Y. Jia, Y.P. Jing, P. Ge, L. Lin and L. Zhang, 2015. Potential association between the protective effects of metformin and catalase in mice with endotoxic liver injury. J. Chin. Pharm. Sci., 24: 399-402.
Direct Link