
Mohammed Dadach
Laboratory of Plant Biodiversity Development and Conservation, Faculty of Natural Sciences and Life, Djillali Liabes University, Sidi Bel Abbes, 22000, Algeria
Zoheir Mehdadi
Laboratory of Plant Biodiversity Development and Conservation, Faculty of Natural Sciences and Life, Djillali Liabes University, Sidi Bel Abbes, 22000, Algeria
Ali Latreche
Laboratory of Plant Biodiversity Development and Conservation, Faculty of Natural Sciences and Life, Djillali Liabes University, Sidi Bel Abbes, 22000, Algeria
Journal of Plant Sciences
Year: 2015 | Volume: 10 | Issue: 4 | Page No.: 151-158







ABSTRACT
Thymus serpyllum L. is a xerophytic perennial herb belongs to the Lamiaceae family commonly used for medicinal purposes. The aim of this study is to determine the effects of water potential on germination. Seeds were germinated under stress of aqueous polyethylene glycol (PEG) solutions mixed to create water stress of -0.03, -0.07, -0.2, -0.5, -1, -1.4 and -1.6 MPa. Distilled water was used as the control (0.0 MPa). A preliminary experiment showed 20°C as the optimum germination temperature for seeds of this species. The decrease of water potential progressively declined Initial Germination Percentage (IGP) and Final Germination Percentage (FGP). The highest germination percentages (90%) were obtained under control conditions (without PEG) and increasing moisture stress progressively inhibited seed germination. Although, germination started to regress significantly at -0.5 MPa and decline completely at -1.6 MPa. However, the Initial Germination Day (IGD) and Mean Time of Germination (MTG) increased with decrease in water potential. We conclude that T. serpyllum is able to act as pioneer plant due to the ability of seeds to germinate under lowest water potential.
PDF Abstract XML References Citation
Received: June 19, 2015;
Accepted: July 29, 2015;
Published: August 26, 2015
How to cite this article
Mohammed Dadach, Zoheir Mehdadi and Ali Latreche, 2015. Effect of Water Stress on Seed Germination of Thymus serpyllum L. from Tessala mount. Journal of Plant Sciences, 10: 151-158.
DOI: 10.3923/jps.2015.151.158
URL: https://scialert.net/abstract/?doi=jps.2015.151.158
DOI: 10.3923/jps.2015.151.158
URL: https://scialert.net/abstract/?doi=jps.2015.151.158
INTRODUCTION
Seed germination is an important and vulnerable stage in the life cycle of terrestrial Angiosperms, determines seedling establishment and plant growth. Seed germination is regulated by the interaction of environmental conditions and the state of physiological readiness (Steckel et al., 2004). Each plant species has a specific range of environmental requirements necessary for germination (Baskin and Baskin, 1998). Success of natural propagation depends mainly on the response of the seeds towards the interference of various external factors. Therefore, seed germination success may reflect upon population size, distribution and abundance (Rojas-Arechiga et al., 1998; Flores and Briones, 2001; Ramirez-Padilla and Valverde, 2005). Indeed, the environmental conditions of the area of species occurrence are essential to determine the seed characteristics and its germination responses. Mainly, the temperature, water availability, soil or substrate type and the rate of gas exchange can promote or inhibit the germination and then influence the seed germination process (Fenner and Thompson, 2005; Cota-Sanchez and Abreu, 2007). All of those factors influence the germination alone or associated among them, so each portion or population of seeds shows different responses to the environment variations.
North Africa has a Mediterranean climate, which is characterized by its seasonality in temperature and precipitation, which leads to a hot drought period in summer and a cool wet period in winter (Joffre et al., 1999) and little rainfall approximately 200-400 mm per year for semi-arid zones ecosystems. This peculiarity of the Mediterranean climate has important implications on plant germination physiology, since dry summer conditions limit water availability and thus germination, while cool winter temperatures also limit germination during the season with high water availability (Rundel, 1996).
A considerable number of medicinal and aromatic plants are locally adapted and considered as native to arid zones of the world (Bannayan et al., 2006). The potential of water tolerance and the economical value of medicinal and aromatic plants make them suitable alternative crops in dry land agro-ecosystems (Koocheki and Nadjafi, 2003).
Thymus is considered a genus, characteristic of the Mediterranean, as representatives of all its sections except Thymus sect. Serpyllum (Mill.) Benth. and sect. Hyphodromi (A. Kern.) Halacsy (subsect. Serpyllastrum villar) founded in the other Regions (Morales, 2002). Thymus serpyllum (Lamiaceae family) is a well-known as wild thyme (Nikolic et al., 2014) and has been extensively used in folk medicine for many years (Aziz and Habib-Ur-Rehman, 2008). The information sources indicate that this plant likes soil of medium acidity (Ellenberg, 1991). Loziene (2002) have noted that this species can live in low alkaline sole and it is found in habitats where there is good lightning.
One methodological approach commonly used to detect water stress as a limiting factor for seed germination is the application of PEG as an osmotic medium (Michel and Kaufmann, 1973). In this study, furnished data about time, rate and synchronicity of seed germination of T. serpyllum under water stress in order to better understand the drought tolerant. Information from this study, provides basic knowledge about germination requirements that can be used to determine tolerance of drought stress and recruitment of this species.
MATERIALS AND METHODS
Plant materiel: Seeds of T. serpyllum were collected in late summer, 2014 from Tessala mount in Algeria (province: Sidi Bel Abbes). This mount is semi-arid with a typical Mediterranean climate (average annual rainfall between 290 and 420 mm, Average monthly temperatures are between 9.4-26.6°C, the minimum average temperature of the coldest month varies between 2.5-3.3°C and the mean maxima of the hottest month rarely exceed 30°C). Seeds for the germination experiments were collected after maturation (harvest station is located in the following geographical coordinates: 35°16’19.19" N; 00°46’02.14" W at 802 m).
Germination experiments: After collection, immature and insect-damaged seeds were removed. Surface seeds were sterilized by soaking in sodium hypochlorite (NaOCl 1%) for 5 min and then thoroughly rinsed with deionised water (Nadjafi et al., 2009). Germination experiments were conducted in darkness in incubators (Memmert type). The seeds were placed in Petri dishes of 9 cm diameter fitted with two layers of Whatman No. 1 filter paper. Each treatment consisted of 4 replicates with 25 seeds. Seeds were germinated, when radical appeared (Zeng et al., 2014), germinated seeds were counted and removed every tow day over a period of 20 days.
Optimization of germination temperature: Since, no information is available on seed germination of T. serpyllum, a preliminary experiment was carried out in order to determine its optimal temperature.
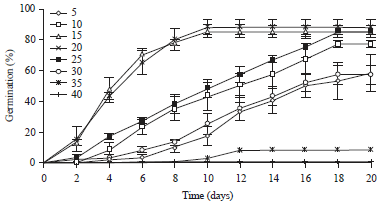 | |
| Fig. 1: | Cumulative germination percentage at different incubation temperatures (°C). Bars represent ±SE (n = 4). The confidence intervals were calculated at the threshold of 5% |
Germination experiments were conducted in incubators set at 5, 10, 15, 20, 25, 30 and 40°C. Distilled water equal to the mean water loss from dishes was added every two days to maintain humidity. We found that the seeds germinated rapidly in distilled water in the range 10-30°C. The optimal temperature for germination was 20°C and the maximum percentage (88%) was achieved after 10 days (Fig. 1). Hence, 20°C was chosen for the study of seed germination responses to water stress in this species.
Effect of water potential: Polyethylene glycol (PEG600) was dissolved in distilled water to make solutions that had the osmotic potentials of -0.03, -0.07, -0.2, -0.5, -1, -1.4 and -1.6 MPa, using the method described by Michel and Kaufmann (1973). Distilled water was used as the control (0.0 MPa). Irrigation with different PEG concentration equal to the mean loss from dishes was added every two days to maintain humidity.
Data analysis: Five germination variables were determined; Final Germination Day (FGD), Initial Germination Percentage (IGP), Final Germination Percentage (FGP), germination kinetics and Mean Time of Germination (MTG).
MTG was calculated as follows:
MTG = Σi (nix di)/N
where, ni is the number of seeds germinated at day i, di is the incubation period in days and N is the total number of germinated seeds (Redondo-Gomez et al., 2007).
Statistical analysis: Data were analysed using SPSS for windows, version 20. Analysis of variance (ANOVA) was carried out to test effects of the main factors on the final germination percentage and mean time of germination. Duncan test was used to estimate least significant range between means.
RESULTS
Effect of water potential on FGP: The decrease of water potential progressively decreases germination (Fig. 2). Thymus serpyllum seeds was significantly (p<0.05) inhibited by PEG concentration higher than -0.2 MPa (Table 1). The highest FGP was founded in distilled water (90%), followed by -0.03 MPa (83%).
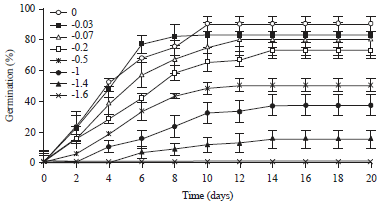 | |
| Fig. 2: | Cumulative germination percentage in response to water potential (MPa). Bars represent ±SE (n = 4). The confidence intervals were calculated at the threshold of 5% |
| Table 1: | Germination characteristic variables of Thymus serpyllum seeds in response to water potential (Mean±SE, n = 4) |
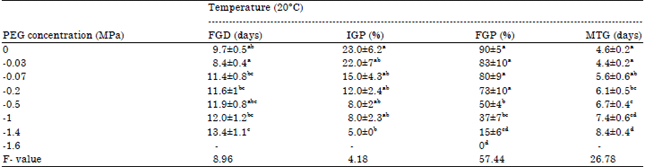 | |
Different lowercase letters (column) show significant differences between the averages (p<0.05), according to Duncan multiple | |
Higher PEG concentration (-1.4 MPa) showed substantial reduction of FGP (15%), germination was completely inhibited at a concentration of -1.6 MPa (Fig. 1).
Effect of water potential on FGD, IGP and MTG: Analysis of variance revealed a significant effect of PEG on the parameters FGD, IGP and MTG (Table 1). At -0.03 MPa it is found the quickest time of FGD (8.4 days). This time started to prolong with the application of stress and the slowest time is registered at -1.4 MPa (13 days). The IGP regressed with increasing water stress with the lowest percentage at concentration of -1.4 MPa (5%). However, MTG increased with PEG concentration and we recorded (4.6 days) in the absence of stress and (8.4 days) at -1.4 MPa.
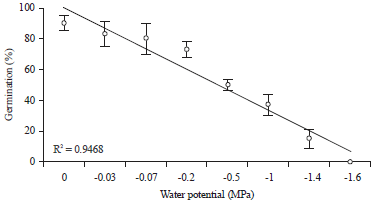
Polynomial analysis: Linear regression analysis was used to determine the relationships between the two parameters (FGP/MTG) and water stress at temperature of 20°C. There is a strong relationship between (FGP/MTG) and PEG concentration with a regression coefficient of 0.94 for the first (Fig. 3) and 0.96 for the second (Fig. 4).
Relationship between FGP and MTG at various water potential concentrations: Nevertheless, a clear distinction between the effect of PEG concentration on the FGP and MTG is evident, when examining, Fig. 5. Actually, water stress affects significantly germination percentage (estimated by the increase of MTG).
 | |
| Fig. 3: | Regression plot for Final Germination Percentage (FGP) in response to water potential. Bars represent ±SE (n = 4). The confidence intervals were calculated at the threshold of 5% |
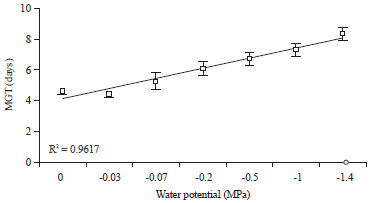 | |
| Fig. 4: | Regression plot for Mean Time to Germination (MTG) in response to water potential. Bars represent ±SE (n = 4). The confidence intervals were calculated at the threshold of 5% |
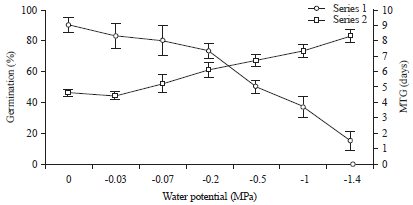 | |
| Fig. 5: | Effect of various water potential concentrations FGP and MTG at 20°C. Bars represent ±SE (n = 4). The confidence intervals were calculated at the threshold of 5% |
DISCUSSION
The negative effect of water stress on germination was reported by many authors for severals plants species (Amara et al., 2013; Rios-Rojas et al., 2014; Zhou et al., 2015). For this species, water soil has more complicated effects on germination than temperature because water is the initial factor for seed imbibitions and germination; it is directly and indirectly involved in subsequent germination metabolic stages (Cavalcante and de Perez, 1995) in particularly under arid and semi arid environment. Our results join those of Abbad et al. (2011), who reported that seeds of Thymus maroccanus and Thymus broussonetii are positively affected by the water potential and seeds germinate better in the absence of stress.
The decrease in germination with decrease of water potential confirms the studies of Smith (1991) and Bafeel (2014) that used sodium chloride solution as the osmoticum. At a concentration of -1 MPa it is found that the FGP drops below 50%, this could be explained by the idea that osmotic potential of 0.5 MPa is a border line for starting the reduction of germination features. The reduction of FGP may be due to alteration of enzymes and hormones found in the seed (Botia et al., 1998). It could also be a deficit of hydration of seeds due to high osmotic potential causing inhibition of the mechanisms leading to the output of the radicle out of the integuments and therefore delay seed germination (Gill et al., 2003). This osmo-regulation mechanism affects the later stages of a growth with slowdown due to the lack and/or unavailability of carbohydrates (Benjelloun et al., 2013).
The water stress thus decrease both FGP and speed of germination, which may be due to the seeds being unable to imbibe sufficient water from their surroundings, when as water availability becomes increasingly restricted. Such delays had been observed for Thymus koteschanus and Thymus daenensis seeds (Bagheri et al., 2011).
The seed germination of T. serpyllum under Mediterranean climate conditions starts in autumn, when precipitation usually initiates and the temperature is adequate (Benvenuti et al., 2001). However, the global change effects on the Mediterranean climate is likely to provide more frequent and longer drought periods, increases of the temperature and changes in precipitation patterns in arid regions (Osborne et al., 2000). This problem can negatively affect sexual reproduction in of many Mediterranean species (Ramirez et al., 2008). Indeed, the species germination requires the availability of resource, essentially high soil moisture content. That’s why T. serpyllum probably uses vegetative reproduction as a space colonization strategy.
CONCLUSION
These experiments, under water stress, revealed that germination of T. serpyllum is affected by water availability with a limit of about -1.6 MPa under which germination is inhibited. Therefore, germination may occur when suitable moisture conditions are present. Undoubtedly, future projections of climate change will impact in the population dynamics of this species by influencing seed germination and consequently their potential for natural regeneration.
ACKNOWLEDGMENT
We thank Mr. Sofiane Bouazza for his help in statistical analysis. Pr Belhassaini Hachemi is acknowledged for his help in laboratory study.
REFERENCES
- Abbad, A., R. Belaqziz, K. Bekkouche and M. Markouk, 2011. Influence of temperature and water potential on laboratory germination of two Moroccan endemic thymes: Thymus maroccanus Ball. and Thymus broussonetii Boiss. Afr. J. Agric. Res., 6: 4740-4745.
Direct Link - Amara, I., M. Capellades, M.D. Ludevid, M. Pages and A. Goday, 2013. Enhanced water stress tolerance of transgenic maize plants over-expressing LEA Rab28 gene. J. Plant Physiol., 170: 864-873.
CrossRefDirect Link - Bafeel, S.O., 2014. Physiological parameters of salt tolerance during germination and seedling growth of Sorghum bicolor cultivars of the same subtropical origin. Saudi J. Biol. Sci., 21: 300-304.
CrossRefDirect Link - Bagheri, M., H. Yeganeh, E.Z. Esfahan and M.B. Savadroodbari, 2011. Effects of water stress on seed germination of Thymus koteschanus Boiss. and Hohen and Thymus daenensis Celak. Middle-East J. Sci. Res., 8: 726-731.
Direct Link - Bannayan, M., F. Nadjafi, M. Rastgoo and L. Tabrizi, 2006. Germination properties of some wild medicinal plants from Iran. Seed Technol., 28: 80-86.
Direct Link - Benvenuti, S., L. Andolfi and M. Macchia, 2001. Light and temperature dependence for germination and emergence of white horehound (Marrubium vulgare L.) seeds. Seed Technol., 23: 138-144.
Direct Link - Botia, P., M. Carvajal, A. Cerda and V. Martinez, 1998. Response of eight Cucumis melo cultivars to salinity during germination and early vegetative growth. Agronomie, 18: 503-513.
Direct Link - Cota-Sanchez, J.H. and D.D. Abreu, 2007. Vivipary and offspring survival in the epiphytic cactus Epiphyllum phyllanthus (Cactaceae). J. Exp. Bot., 58: 3865-3873.
Direct Link - Flores, J. and O. Briones, 2001. Plant life-form and germination in a Mexican inter-tropical desert: Effects of soil water potential and temperature. J. Arid Environ., 47: 485-497.
CrossRefDirect Link - Gill, P.K., A.D. Sharma, P. Singh and S.S. Bhullar, 2003. Changes in germination, growth and soluble sugar contents of Sorghum bicolor (L.) Moench seeds under various abiotic stresses. Plant Growth Regul., 40: 157-162.
CrossRefDirect Link - Joffre, R., S. Rambal and J.P. Ratte, 1999. The dehesa system of southern Spain and Portugal as a natural ecosystem mimic. Agroforestry Syst., 45: 57-79.
CrossRefDirect Link - Michel, B.E. and M.R. Kaufmann, 1973. The osmotic potential of polyethylene glycol 6000. Plant Physiol., 51: 914-916.
CrossRefPubMedDirect Link - Nadjafi, F., L. Tabrizi, J. Shabahang and A.M.M. Damghani, 2009. Cardinal germination temperatures of some medicinal plant species. Seed Technol., 31: 156-163.
Direct Link - Nikolic, M., J. Glamoclija, I.C.F.R. Ferreira, R.C. Calhelha and A. Fernandes et al., 2014. Chemical composition, antimicrobial, antioxidant and antitumor activity of Thymus serpyllum L., Thymus algeriensis Boiss. and Reut and Thymus vulgaris L. essential oils. Ind. Crops Prod., 52: 183-190.
CrossRefDirect Link - Osborne, C.P., P.L. Mitchell, J.E. Sheehy and F.I. Woodward, 2000. Modelling the recent historical impacts of atmospheric CO2 and climate change on Mediterranean vegetation. Global Change Biol., 6: 445-458.
CrossRefDirect Link - Ramirez, D.A., F. Valladares, F. Domingo and J. Bellot, 2008. Seasonal water-use efficiency and chlorophyll fluorescence response in alpha grass (Stipa tenacissima L.) is affected by tussock size. Photosynthetica, 46: 222-231.
CrossRefDirect Link - Ramirez-Padilla, C.A. and T. Valverde, 2005. Germination responses of three congeneric cactus species (Neobuxbaumia) with differing degrees of rarity. J. Arid Environ., 61: 333-343.
CrossRefDirect Link - Redondo-Gomez, S., E. Mateos-Naranjo, C. Wharmby, C.J. Luque, J.M. Castillo, T. Luque, M.F. Mohamed, A.J. Davy and M.E. Figueroa, 2007. Bracteoles affect germination and seedling establishment in a Mediterranean population of Atriplex portulacoides. Aquat. Bot., 86: 93-96.
CrossRefDirect Link - Rios-Rojas, L., F. Tapia and L.A. Gurovich, 2014. Electrophysiological assessment of water stress in fruit-bearing woody plants. J. Plant Physiol., 171: 799-806.
CrossRefDirect Link - Rojas-Arechiga, M., C. Vazquez-Yanes and A. Orozco-Segovia, 1998. Seed response to temperature of Mexican cacti species from two life forms: An ecophysiological interpretation. Plant Ecol., 135: 207-214.
CrossRefDirect Link - Rundel, P.W., 1996. Monocotyledonous geophytes in the California flora. Madrono, 43: 355-368.
Direct Link - Steckel, L.E., C. Sprague, E.W. Stoller and L. Wax, 2004. Temperature effects on germination of nine Amaranthus species. Weed Sci., 52: 217-221.
CrossRefDirect Link - Zeng, Y.J., Y.R. Wang, C.C. Baskin and J.M. Baskin, 2014. Testing seed germination responses to water and salinity stresses to gain insight on suitable microhabitats for restoration of cold desert shrubs. J. Arid Environ., 100: 89-92.
CrossRefDirect Link - Zhou, S., Y.Y. Han, Y. Chen, X. Kong and W. Wang, 2015. The involvement of expansins in response to water stress during leaf development in wheat. J. Plant Physiol., 183: 64-74.
CrossRefDirect Link