
Fisseha Worede
Amhara Regional Agricultural Research Institute, Sirinka Research Center, Ethiopia
Tanee Sreewongchai
Department of Agronomy, Faculty of Agriculture, Kasetsart University, Bangkok, Thailand
Chalermpol Phumichai
Department of Agronomy, Faculty of Agriculture, Kasetsart University, Bangkok, Thailand
Prapa Sripichitt
Department of Agronomy, Faculty of Agriculture, Kasetsart University, Bangkok, Thailand
Journal of Plant Sciences
Year: 2014 | Volume: 9 | Issue: 1 | Page No.: 14-24







ABSTRACT
The availability of access to diverse genetic material is important to be successful in any plant breeding effort. An investigation was undertaken to assess genetic diversity of 24 known rice genotypes based on17 morpho-agronomic traits by using multivariate analyses: hierarchical cluster and Principal Component (PC) analyses. Cluster analysis separated the genotypes into two groups which are in contrast for flowering, plant types, biomass, grain yield, seed width, seed length and seed weight. The first three PCs explained 73.5% of the total variability. Days to flowering, plant height, culm length, panicle length, biomass, seed length and shape were the important traitsin differentiating the genotypes. The hierarchical cluster and PC analyses were in agreement in grouping the genotypes. Parental combinations from the two clusters with higher value of Euclidean distance could be used for genetic improvement. Generally KOH1 is the most diverged genotype from others. Thus crossing of this genotype with Azucena, CH1, KDML105 and SPR1 may result in heterotic expression in the F1 and substantial variability in the subsequent segregating population.
PDF Abstract XML References Citation
Received: August 06, 2013;
Accepted: January 06, 2014;
Published: February 27, 2014
How to cite this article
Fisseha Worede, Tanee Sreewongchai, Chalermpol Phumichai and Prapa Sripichitt, 2014. Multivariate Analysis of Genetic Diversity among some Rice Genotypes Using Morpho-agronomic Traits. Journal of Plant Sciences, 9: 14-24.
DOI: 10.3923/jps.2014.14.24
URL: https://scialert.net/abstract/?doi=jps.2014.14.24
DOI: 10.3923/jps.2014.14.24
URL: https://scialert.net/abstract/?doi=jps.2014.14.24
INTRODUCTION
More than half of the population of the world depends on rice as the main source of food. Rice is one of the world’s most important food crops in terms of its production and area coverage. About 11% of the world’s cultivated land is devoted for its production. Among the continents of the world, Asia is the first both in production and consumption of rice. Most of the Asians get 35-75% of their calories from rice (Khush, 2005). In the year 2011 about 164.1million ha of land was used for rice production worldwide. In the same year nearly equal amount of land was allocated for rice in the content of Africa and in Thailand (FAO, 2013).
Genetic diversity has a multitude of importance. It is considered as a means of survival and adaptation to changing environments (Gao, 2003; Rao and Hodgkin, 2002); it has a role in collection and conservation of germplasm and crop improvement (Duran et al., 2009). Moreover, the information helps to study heterosis as genetic diversity between parents is generally related to the magnitude of heterosis (Virk et al., 2003). This shows the necessity of divergent parents in any crossing program in order to be benefited from the hybrid vigor in the F1 and from the variability created in the subsequent segregating population. Whether the objective is hybrid or pure-line variety development, the availability of and access to diverse genetic material is important to be successful in any plant breeding effort (Hoisington et al., 1999; Naik et al., 2006; Maxted et al., 2002; Rohman et al., 2004).
Evaluation of genetic resources for various agronomic traits and the assessment of the amount of genetic variation within them is useful to allow more efficient genetic improvement (Haussmann et al., 2004). In order to further improve the already adapted crop varieties for traits of agronomic importance, crossing with other parents with proved agronomic superiority is necessary. This calls for the assessment of genetic diversity and identification of parental lines for hybridization or crossing. The present investigation was undertaken with the objective to assess genetic diversity of some known rice genotypes by using morpho-agronomic traits and to identify the most important character in differentiating the genotypes.
MATERIALS AND METHODS
Experimental materials and place of study: In the genetic diversity study, a total of 24 rice genotypes were used. The materials include 13 indica, four japonica, four New Plant Types, one tropical japonica and two NERICA genotypes (Table 1). NERICA signifies New Rice for Africa; genotypes developed by interspecific hybridization of O. glaberrima and O. sativa (Samado et al., 2008). The trial was conducted at Kasetsart University, Bangkhen Campus, Bangkok, Thailand during 2012 rainy season.
Experimental design and data collection: The treatments were arranged in a randomized complete block design with three replications. A spacing of 20 cm between genotypes and between plants was left.
| Table 1: | Description of the 24 rice genotypes used for the diversity analysis using 17 morpho-agronomic traits |
 | |
Fertilization and insecticide application were done uniformly to all plots. The plants were protected from the possible damage of birds and rats by wire mesh. Data for 17 morphological and agronomic traits were collected from each plot using the Standard Evaluation System for rice (IRRI, 2002). Paddy-rice seed width and length were measured as averages of 15 seeds by using digital Rice Grain Analyzer. The traits studied include days to flowering, No. of total tillers, number of productive tillers, plant height (cm), culm length (cm), flag-leaf length (cm), panicle length (cm), number of total spikelets, number of fertile spikelets, 100-seed weight (g), biomass (g), yield per plant, harvest index (%), seed width (mm), seed length (mm), seed shape (length, width ratio) and spikelet fertility (%).
Data analyses: Analysis of Variance (ANOVA) was computed by using CropStat7.2 and the mean values of traits were used for further analysis. The mean values were then standardized to a mean of zero and variance of unity before cluster analysis to remove the biases due to differences in the scale of measurement. PAST 1.93 (Palaeontological Statistics; Hammer et al., 2001) computer software was used for phenotypic correlation, cluster and principal component analyses.
RESULTS
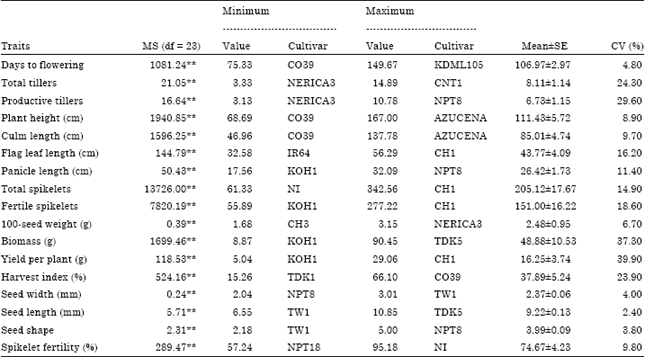
Analysis of variance: The univariate ANOVA showed significant (p<0.01) variation among the rice genotypes for all the morpho-agronomic traits considered (Table 2). The significance signifies the possibility of using all traits for further analysis. From the result it was observed that CO39 was the earliest (75.3 days) while KDML105 was the latest (149.67 days) genotypes to flower. Similarly, CO39 and Azucena were the shortest (68.69 cm) and the tallest (167 cm) genotypes, respectively.
| Table 2: | Minimum and maximum mean values and overall means, standard errors and coefficients of variation of 17 morpho-agronomic traits of 24 rice genotypes |
 | |
| **: Significant at 0.01 probability level | |
The analysis also showed that KOH1 was the lowest (5.04 g) whereas, CH1 was the highest (29.06 g) in yield among the genotypes. Also, NPT18 and NI were the least (57.24%) and the most (95.18%) fertile genotypes identified, respectively.
Phenotypic correlation: In order to assess traits association, phenotypic correlation analysis was done and the result is depicted in Table 3. Generally the result showed high order of correlation between most of the traits under study. Days to flowering was highly and significantly correlated with plant height (0.883), culm length (0.893), biomass (0.772) and harvest index (-0.732). Similarly, yield per plant was highly and significantly correlated with total tillers (0.541), plant height (0.525), panicle length (0.649), total spikelets (0.585), filled spikelets (0.539), biomass (0.716), seed length (0.606) and seed shape (0.622) towards the positive direction. Also, strong and significant association of harvest index was detected with days to flowering (-0.732), plant height (-0.603), culm length (-0.601), total spikelets (-0.455), biomass (-0.705) and spikelet fertility (0.710). Spikelet fertility was negatively correlated to all traits under study except harvest index.
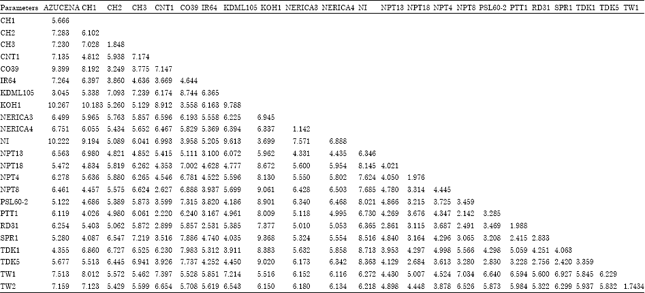
Euclidean distance: Euclidean distance matrix was produced by assuming 276 total possible pairwise combinations of the 24 rice genotypes (Table 4). The distance coefficients ranged from 1.142 for NERICA3-NERICA4 to 10.267 for KOH1-Azucena pairwise cultivar combinations with a mean of 5.172. Also TW1-TW2 (1.743), CH2-CH3 (1.848) and NPT4-NPT18 (1.976) were the next smaller pairwise Euclidean distances. Similarly, the next higher distance values were that of KOH1 with CH1 (10.183), KDML105 (9.788) and SPR1 (9.368). KOH1 is generally the most diverged genotype from others with higher mean Euclidean distance of 7.426 while RD31 was the least with mean Euclidean distance of 4.41.
Hierarchical cluster analysis: Hierarchical clustering was attempted by using paired group algorithm with different distance measures like Gower, Euclidean, Mahalanobis and Manhattan. The result showed that Gower, Euclidean and Manhattan distance measures yielded similar dendrogram topology and similar cluster membership of the rice genotypes; however, Mahalanobis distance measure yielded different dendrogram topology which was characterized by chaining of genotypes. The dendrogram of the morpho-agronomic traits grouped the genotypes into two clusters with additional subgroups in each groups. Group I was composed of 17 genotypes and group II included 7 genotypes. Group I was made up of miscellaneous type of genotypes as it was composed of 10 indica, four NPT, one tropical japonica and two NERICA types. This group was further divided into four subgroups: Subgroup1 contained the two NERICAs; subgroup 2 was made up of the four NPTs and seven indica types; subgroup 3 contained a distinct cultivar CH1, this variety was characterized by longer flag-leaf, higher number of spikeletes and higher yield and subgroup 4 included KDML105, Azucena and TDK1. Members of this subgroup were late maturing and tall plant types. However, group II contained four japonica and three indica types. This group also further divided in to three subgroups: Subgroup 1 contained TW1 and TW2 (japonica types); subgroup 2 was made up of CH2, CH3 and CO39 (indica types) and subgroup 3 was composed of NI and KOH1, japonica types (Fig. 1). ANOVA was run to declare whether any of the mean differences of the traits between the two groups were significant. Accordingly, the result showed significant (p<0.01) differences for most of the traits under study except for traits related to tillers and spikeletes and harvest index.
| Table 3: | Phenotypic correlation coefficients between 17 morpho-agronomic traits in 24 rice genotypes |
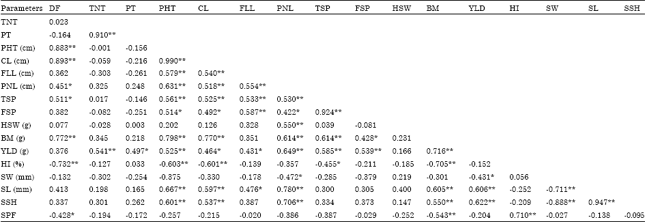 | |
| DF: Days to flowering, TNT: Total No. of tillers, PT: Productive tillers, PHT: Plant height, CL: Culm length, FLL: Flag-leaf length, PNL: Panicle length, TSP: Total spikelets, FSP:Filled spikelets, HSW: Hundred-seed weight, BM: Biomass, YLD: Yield per plant, HI: Harvest index, SW: Seed width, SL: Seed length, SSH: Seed shape, SPF: Spikelet fertility**,*: Significant at 0.01 and 0.05 probability levels, respectively | |
| Table 4: | Pairwise Euclidean distance coefficients for all possible combinations of the 24 genotypes using 17 morpho-agronomic traits |
 | |
 | |
| Fig. 1: | Dendrogram of 24 rice genotypes based on 17 morpho-agronomic traits constructed by means of paired group algorithm and Euclidean distance |
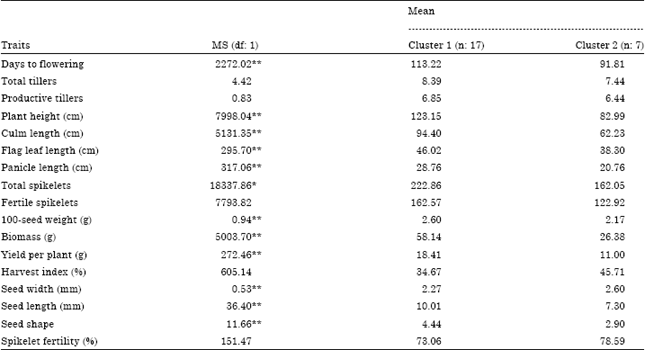
On average, the genotypes in cluster II were significantly earlier (91.8 vs. 113.2 days) and shorter (82.99 vs. 123.15 cm) than that of cluster I. Compared with cluster I, the genotypes in cluster II had significantly lower biomass (26.38 vs. 58.14 g) and grain yield (11 vs. 18.41 g) (Table 5).
Principal component analysis: The result of the principal component analysis (PCA) is depicted in Fig. 2 and Table 6. The objective of principal component analysis is reduction of dimensionality of a data set with a large number of correlated variables or traits (Jolliffe, 2002). PCA was carried out by using 24 genotypes and 17 traits. In the analysis a total of 17 PCs, equals to the number of traits, were extracted. However, the first five PCs with eigen values greater than 1 were retained. The result showed that 89.68% of the variability was explained by the first five Principal Component (PC) axes. Out of the five, the first and the second explained 44.52 and 16.64% of the variation, respectively. Days to flowering, plant height, culm length, panicle length, biomass, seed length and shape were the important traits contributing to the first PC. In the second PC, however, total and productive tillers were important. Similarly, flag-leaf length, harvest index, seed width and spikelet fertility were important in the third axis. While only 100-seed weight was important in the fourth axis. Total and filled spikelets and yield per plant were the important traits contributing to the fifth PC (Table 6). The first axis differentiated genotypes which were late flowering, tall, higher in biomass and grain yield, with slender (narrow and tall) seeds from genotypes which were early flowering, short, low biomass and grain yield with round (wide and short) seeds. The second axis however differentiated genotypes with higher total and productive tillers from those with low tillers. Generally the PC analysis broadly grouped the genotypes based on zones of origin into temperate and tropical categories.
| Table 5: | Mean-squares and cluster means for 17 morpho-agronomic traits of the 24 rice genotypes |
 | |
| **,*: Significant at 0.01 and 0.05 probability levels | |
| Table 6: | Eigenvalues, total variance, cumulative variance and eigenvectors for 17 morpho-agronomic traits in the 24 rice genotypes |
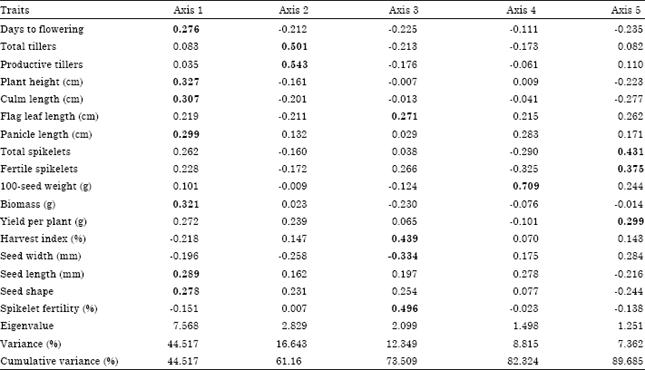 | |
The ones located to left of the two dimensional plane being temperate with the exception of CO39, while those to the right being tropical in zone of origin (Fig. 2).
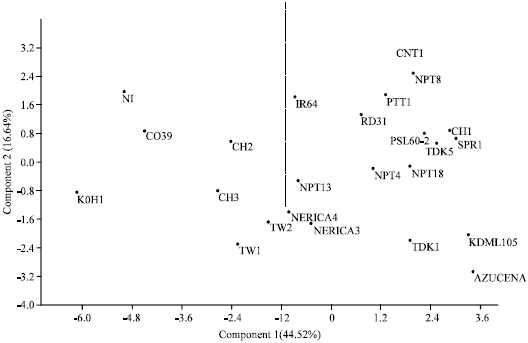 | |
| Fig. 2: | Scattered distribution of the 24 rice genotypes by using 17 morpho-agronomic traits on the first two principal component axes |
DISCUSSION
The significant (p<0.01) variation among the rice genotypes for all the morpho-agronomic traits and the range of values obtained for most of the traits indicates a sizable variability in the genotypes studied for the 17 traits considered. The correlation study showed that late flowering is important in attaining tall plant types, higher biomass and lower harvest index. It also showed that more number of tillers, tall plant types (increased height and panicle length), higher number of spikelets, heavier above ground mass, seed length and shape were the important traits in attaining higher yield per plant in rice. However, early flowering, short plant types (lower height and culm length), lower number of total spikelets, light above ground-mass but higher level of spikelet fertility are important to improve harvest index. Significant correlations of yield and yield-related traits were reported by other workers in inbred lines, hybrids and landrace rice (Chakravorty et al., 2013; Janwan et al., 2013; Seesang et al., 2013).
The pairwise Euclidean distance coefficients estimated in this study ranged from 1.142-10.267 with a mean of 5.172. However, Caldo et al. (1996a) estimated a range of Euclidean distance varying between 2.23 and 16.71 with a mean of 7.55 for 78 improved rice genotypes by using 33 qualitative and quantitative traits. In a separate study of 81 ancestral lines of Philippines modern rice genotypes, Caldo et al. (1996b) computed Euclidean distance estimates ranging from 3.97-17.389 with a mean of 8.80 using 41 traits.The output of hierarchical cluster analysis exhibited similar dendrogram topology and cluster membership of the rice genotypes for Gower, Euclidean and Manhattan distance measures. This confirms the stability of the dendrogram constructed. Generally, the first group was characterized by late flowering, tall plant types, higher biomass and grain yield, slender (narrow and long) and heavier seeds. Unlike the first group, the second group was characterized by early flowering, short plant types, lower biomass and grain yield, round (wide and short) and light weighted-seeds.
Principal Components (PCs) are orthogonal and independent of each other (Mohammadi and Prasanna, 2003); they explain the variability which was not explained by the others. In this study, the total variability was explained by five PCs. This may indicate the contribution of many traits with higher level of correlation to explain the gross diversity. In a study of Caldo et al. (1996b), 75% of the variability was accounted for by seven PCs. In their work, Chakravorty et al. (2013) explained the total variability by six PC axes; Caldo et al. (1996a, b) explained 67% of the variation by 10 PCs. Jolliffe (2002) stated that PCs are ordered so that the first few retain most of the variation present in all of the original variables. In this experiment, the first and the second PCs explained 61.16% of the variability, the first being the most important. Accordingly, the traits included in the first PC especially those with comparatively high loadings are important in separating the genotypes. Generally, the PC analysis showed high level of diversity of the rice genotypes as the entire variation cannot be explained by few principal components. High level of simple sequence repeat (SSR) DNA marker diversity was also reported for the same set of genotypes investigated (Worede et al., 2013).
CONCLUSION
The study showed the presence of considerable level of diversity in the genotypes analyzed and the importance morpho-agronomic traits to study genetic diversity. Days to flowering, plant height, culm length, panicle length, biomass, seed length and shape were the important traits in differentiating the genotypes under study. The hierarchical cluster analysis was in general agreement with the PC analysis in grouping the genotypes into two clusters. Parental combinations from the two clusters with higher value of Euclidean distance (dissimilarity) coefficient could be used for genetic improvement by crossing. Generally KOH1 is the most diverged genotype from others. Thus crossing of this genotype with the likes of Azucena, CH1, KDML105 and SPR1 may result in heterotic expression in the F1 and substantial variability in the subsequent segregating population.
ACKNOWLEDGMENT
The study was supported by Rural Capacity Building and East Africa Agricultural Productivity Projects of the Ethiopian Ministry of Agriculture.
REFERENCES
- Caldo, R.A., L.S. Sebastian and J.E. Hernandez, 1996. Multivariate analysis of phenotypic diversity of Philippine improved rice genotypes. Philipp. J. Crop Sci., 21: 76-85.
Direct Link - Caldo, R.A., L.S. Sebastian and J.E. Hernandez, 1996. Morphology-based genetic diversity analysis of ancestral lines of rice in Philippine rice cultivars. Philipp. J. Crop Sci., 21: 86-92.
Direct Link - Chakravorty, A., P.D. Ghosh and P.K. Sahu, 2013. Multivariate analysis of phenotypic diversity of landraces of rice of West Bengal. Am. J. Exp. Agric., 3: 110-123.
CrossRefDirect Link - Duran, C., N. Appleby, D. Edwards and J. Batley, 2009. Molecular genetic markers: Discovery, applications, data storage and visualisation. Curr. Bioinform., 4: 16-27.
CrossRef - Gao, L.Z., 2003. The conservation of Chinese rice biodiversity: genetic erosion, ethnobotany and prospects. Genet. Resour. Crop Evol., 50: 17-32.
CrossRefDirect Link - Hammer, O., D.A.T. Harper and P.D. Ryan, 2001. PAST: Paleontological statistics software package for education and data analysis. Paleontol. Electron., 4: 1-9.
Direct Link - Haussmann, B.I.G., H.K. Parzies, T. Presterl, Z. Susic and T. Miedaner, 2004. Plant genetic resources in crop improvement. Plant Genet. Resour., 2: 3-21.
Direct Link - Hoisington, D., M. Khairallah, T. Reeves, J.M. Ribaut, B. Skovmand, S. Taba and M. Warburton, 1999. Plant genetic resources: What can they contribute toward increased crop productivity? Proc. Natl. Acad. Sci. USA., 96: 5937-5943.
CrossRefDirect Link - IRRI, 2002. Standard Evaluation System for Rice (SES). International Rice Research Institute, Manila, Philippines, Pages: 56.
Direct Link - Janwan, M., T. Sreewongchai and P. Sripichitt, 2013. Rice breeding for high yield by advanced single seed descent method of selection. J. Plant Sci., 8: 24-30.
CrossRefDirect Link - Khush, G.S., 2005. What it will take to feed 5.0 billion rice consumers in 2030. Plant Mol. Biol., 59: 1-6.
CrossRef - Maxted, N., L. Guarino, L. Myer and E.A. Chiwona, 2002. Towards a methodology for on-farm conservation of plant genetic resources. Genet. Resour. Crop Evol., 49: 31-46.
CrossRefDirect Link - Mohammadi, S.A. and B.M. Prasanna, 2003. Analysis of genetic diversity in crop plants-salient statistical tools and considerations. Crop Sci., 43: 1235-1248.
CrossRefDirect Link - Naik, D., A. Sao, A.K. Sarawgi and P. Singh, 2006. Genetic divergence studies in some indigenous scented rice (Oryza sativa L.) accessions of central India. Asian J. Plant Sci., 5: 197-200.
CrossRefDirect Link - Rao, V.R. and T. Hodgkin, 2002. Genetic diversity and conservation and utilization of plant genetic resources. Plant Cell Tissue Org. Cult., 68: 1-19.
Direct Link - Rohman, M.M., M.A. Hakim, N.A. Sultana, M.E. Kabir, M. Hasanuzzaman and M. Ali, 2004. Genetic divergence analysis in sorghum (Sorghum bicolor L.). Asian J. Plant Sci., 3: 211-214.
CrossRefDirect Link - Seesang, J., P. Sripicchitt, P. Somchit and T. Sreewongchai, 2013. Genotypic correlation and path coefficient for some agronomic traits of hybrid and inbred rice (Oryza sativa L.) cultivars. Asian J. Crop Sci., 5: 319-324.
CrossRef - Virk, P.S., G.S. Khush and S.S. Virmani, 2003. Breeding Strategies to Enhance Heterosis in Rice. In: Hybrid Rice for Food Security, Poverty Alleviation and Environmental Protection, Virmani S.S., C.X. Mao and B. Hardy (Eds.). Int. Rice Res. Inst., Los Banos, Philippines, ISBN: 971-22-0188-0, pp: 21-29.