
Arvind Soni
Plant Genomics, Division of Molecular Biology and Genetic Engineering, National Botanical Research Institute, Lucknow, India
Alka Niveden
Laboratory of Bryology, Division of Biodiversity, National Botanical Research Institute, Lucknow, India
Virendra Nath
Laboratory of Bryology, Division of Biodiversity, National Botanical Research Institute, Lucknow, India
Anil Kumar
Plant Genomics, Division of Molecular Biology and Genetic Engineering, National Botanical Research Institute, Lucknow, India
Journal of Plant Sciences
Year: 2014 | Volume: 9 | Issue: 1 | Page No.: 1-13







ABSTRACT
Random amplified polymorphic DNA markers were used to determine the altitudinal variation within and between Plagiochasma appendicualtum collected from different altitude of Western Himalaya especially from Mussoorie region of India. Findings of UPGMA cluster analysis and band frequency of all the nine accessions were separated according to their altitudes supporting to their morphological differences as well. Gene flow and spore dispersal plays an important role in the polymorphism. Gene flow within P. appendiculatum growing on same altitude is very high as compared to accessions collected from different gradient of altitudes i.e., the genotypes collected from same altitude showing not so much polymorphism compared to different altitude. It has been concluded that the RAPD markers would be useful to characterize the altitudinal variation between different accessions of P. appendiculatum and may be also valuable to other bryophytes collected from various environmental condition.
PDF Abstract XML References Citation
Received: November 27, 2013;
Accepted: December 21, 2013;
Published: February 27, 2014
How to cite this article
Arvind Soni, Alka Niveden, Virendra Nath and Anil Kumar, 2014. Uses of Random Amplified Polymorphic DNA (RAPD) Markers in the Altitudinal
Diversity of Plagiochasma appendicualtum. Journal of Plant Sciences, 9: 1-13.
DOI: 10.3923/jps.2014.1.13
URL: https://scialert.net/abstract/?doi=jps.2014.1.13
DOI: 10.3923/jps.2014.1.13
URL: https://scialert.net/abstract/?doi=jps.2014.1.13
INTRODUCTION
Bryophytes are the diverse group of land plants after the flowering plants (Mishler, 2001) but due to complexity in their identification and lack of literature from tropical areas, they have rarely been included in biodiversity analysis (Pharo et al., 1999). They are found from the tropics to the polar regions, from sea level to mountain summits and are principle candidates for latitudinal and altitudinal revision (Andrew et al., 2003). Several evocative studies of bryophytes and their altitudinal zonation have been reported in South America, in Puerto Rico along a transect of 200-1075 m by Fulford et al. (1971), in the Sierra Nevada de Santa Marta in Colombia (Van Reenen and Gradstein, 1983, 1984). Also in Northeastern Peru (Gradstein and Frahm, 1987), Bolivia, Peru and Columbia (Kessler, 2000), on Mt. Kinabulu, in the eastern part of Borneo (Frahm, 1990) and in New Zealand (Frahm and Ohlemuller, 2001; Pfeiffer, 2003) as well as in Africa on Mount Kilimanjaro (Pocs, 1994).
Mussoorie is a city about 30 km from Dehradun located in Dehradun district, Indian state of Uttarakhand. This hill station, situated in the foothills of the western Himalaya ranges, is also known as the Queen of the Hills. Mussoorie, with its green hills and varied flora and fauna, is notable for its unique geographical location with varied topography and associated altitudinal diversity supporting unique assemblage of biodiversity. Except for the coast and backwaters, almost all habitats in Western Himalaya, from low altitude to the high altitude grasslands and shoal forests, occur in Mussoorie, which forms the major part of the Himalaya Biosphere Reserve.
Plagiochasma appendiculatum is one of the important liverwort belongs to the order Marchantiales under family Aytoniaceae. Lehman and Lindenberg (1832) first described the species. Plagiochasma is a thalloid liverwort represented by 30 species (Bischler, 1978), but in India only 10 species have been reported, viz., Plagiochasma appendiculatum Lehm. et Lindb, Plagiochasma articulatum Kash., Plagiochasma bicornutum Steph., Plagiochasma cordatum Lehm. et Lindb, Plagiochasma cordotii Steph., Plagiochasma intermedium L. et Gott., Plagiochasma martensii Steph., Plagiochasma nepalensis Steph., Plagiochasma pauriana Udar et Chandra and Plagiochasma quadricornutum Steph. (Parihar et al., 1994). Out of these taxa P. appendiculatum abundantly grows in Mussoorie and also on other parts of India as well.
It is a monoecious plant and usually grows in moist places, on rocks surface, soil covered rocks, walls of old buildings and show extra ordinary regeneration (Mahabale and Bhate, 1945). Besides this, P. appendiculatum also represents the maximum xerophytic habitat and can grow on comparatively naked and exposed rocks (Kachroo, 1954). Ghate and Chaphekar (2000) proved that this taxon could be used as a biotest for water quality assessment. P. appendiculatum is significant taxon which possesses antimicrobial property. Banerjee (2000), Kumar et al. (2000) and Singh et al. (2006) stated that in India, it is used by Gaddi tribes in Himachal Pradesh for the treatment of cuts, wounds and burns. Genetic variation of P. appendicualtum collected from different geographical conditions have been reported by using RAPD markers (Soni et al., 2009). Under the same study, RAPD markers were also stated the genetic diversity of P. appendicualtum within and between populations.
Random amplified polymorphic DNA markers (RAPD) can be widely used as DNA fingerprinting techniques (Williams et al., 1990, 1993) that has been utilized in bryophytes to survey population genetic structural, dispersal of spores, phylogeographic patterns and species relationship (Skotnicki et al., 2000; Scotnicki et al., 2001; Freitas and Brehm, 2001; Boisselier-Dubayle and Bischler, 1994; Boisselier-Dubayle et al., 1995). There were several reports related to RAPD genetic variation of various species of Antarctic moss (Selkirk et al., 1997; Skotnicki et al., 1997, 1998a, b).
P. appendiculatum is widely distributed in western, eastern Himalayas, central India and south India and generally growing upto an altitude of 8000 ft from sea level. This species in known from the east part of the central and south African continent Eritrea, Ethiopia, Kenya, Tanzania to Rhodesia, Zimbabwe and South Africa (Perold, 1999; Wigginton, 2002). In Asia, it is widespread ranging from the southwest of the Arabian Peninsula and Socotra Islant (Frey and Kurschner, 1988) to the southern part of the Himalayas, Formosa, Philippines and Celebes (Bischler, 1979). Due to occurrence of such variable altitudinal range, present study was planned out to determine altitudinal variation in P. appendiculatum through random amplified polymorphic DNA marker.
MATERIALS AND METHODS
Field sampling: The samples of P. appendiculatum were collected from various localities of Mussoorie e.g., Library road, Camel’s back road, Kempty fall, Wood stock collage, Lal-Tibbs and Company garden (Fig. 1). Voucher specimens have been deposited at NBRI Bryophyte Herbarium, Lucknow, India (Table 1). Each specimen has been identified by their characteristic features through literature and authentic specimens available in the NBRI Herbarium.
Plant identification: Thalli of Plagiochasma appendiculatum is characterized by large, purplish green patches, thick, 20 mm long and 5 mm wide, dichotomously branched and occasionally with adventitious shoots.
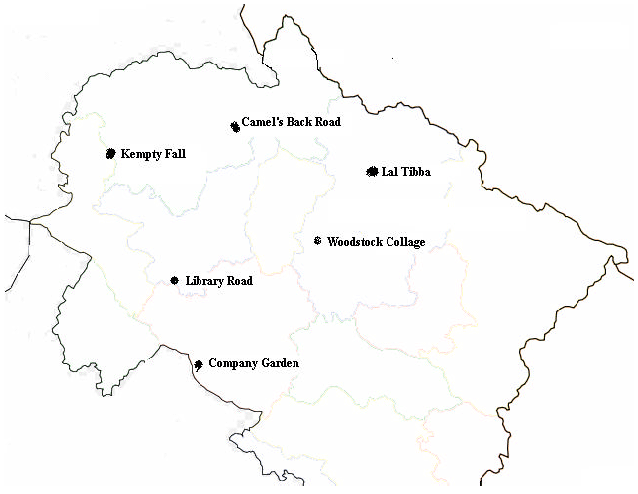 | |
| Fig. 1: | Collection sites of P. appendiculatum from various altitude of Mussoriee (Western Himalaya) India |
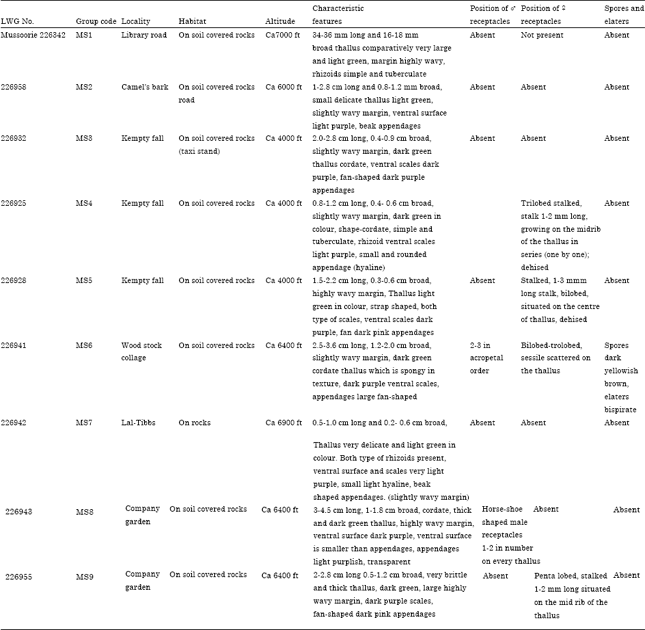
| Table 1: | Genotype collection of P. appendiculatum from various localities of mussoorie, growing at different altitude |
 | |
Lobes oblong, dorsal surface smooth margins undulate. Midrib distinct passing into lamina, ventral surface purple. Scales in one row on each side of the midrib purple, broadly lunate, body with 1 or 2 appendages reaches half way to the margins, appendages large, usually hyaline, fan shaped, entire and occasionally purple. Male receptacle horse-shoe shaped sometimes scattered on the thallus or in acropetal order. Female receptacle sessile or stalked, usually with 2-5 lobes, situated on the thallus in row or scattered or some time at the basal part of the plant. Spores yellowish brown and elaters bispirate. This species is mainly separated from the other species on the basis of its scale structure that has typical from shaped on broad lunate appendages (Table 2).
| Table 2: | Morphological data of Plagiochasma appendiculatum collected from geographical localities of Mussoorie (India) |
 | |
Plant DNA preparation: Fresh and matured thallii of P. appendiculatum were used to isolate the genomic DNA. All samples were carefully washed and checked under the microscope to be sure that no possible contaminations (microalgae, fungi) were left. DNA was extracted from 1 g of plant material with some modification of several standard protocols (Doyle and Doyle, 1990; Soni and Kumar, 2009). Fresh material was crushed in liquid nitrogen and mixed with modified CTAB extraction buffer. Add 1% α-mercaptoethanol, 2% proteinase K (20 mg mL-1) and 100 mg PVP (Sigma) and then incubated in a water bath at 68°C for 5 h, add Chloroform: Isoamyl alcohol (24:1) to this mixture, followed by centrifugation for 10 min at 12,000 rpm. The supernatant was transferred to a corex tube, add 0.7 Vol. iso-propanol, mixed well, store in -20°C for 5 h. to precipitate the DNA. Centrifuge 10 min at 12,000 rpm. Supernatant was discarded and the pellet washed with a solution of 70% ethanol. Tubes were incubated at 37°C for 15 min and resuspended the pellet in TE buffer. Purify the DNA followed by phenol/chloroform method, then precipitate and washing as before. Finally, the aqueous phase was discarded and the pellet was dried for 15 min in incubator at 37°C. Resuspended the pellet in 100 μL TE buffer. This procedure recovered at least 800 ng μL-1 genomic DNA which is good quality for RAPD analysis.
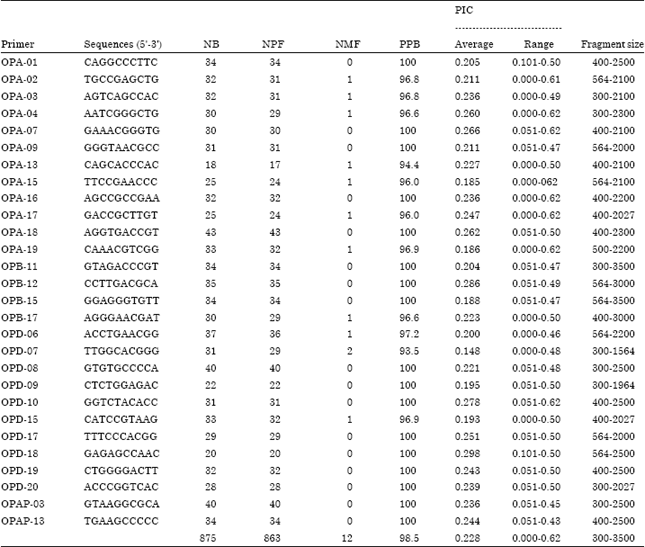
Primers and PCR: Decamer primers from OPERON Technologies (USA) were screened on individual representative of the populations under study. Many of the primer produced either complex banding pattern of non-reproducible and inconsistent amplification products. Hence, only 45 primers scored good result out of 60 primers used for the subsequent analysis (Table 3). Reproducibility of bands was assessed by replicating extraction of DNA and amplifications of selected samples. Polymorase Chain Reaction (PCR) was carried out in 20 μL volumes using DNA, dNTPs (2 mM of each of four nucleotides: Fermentas), 10X Taq Buffer, 5 pmol primer, 1 unit Taq DNA polymerase (Bangalore Genei). PCR conditions were initiated at 92°C followed by 44 cycles of denaturation at 92°C for 1 min, annealing at 36°C for 1 min and extension at 72°C for 1:30 min, followed by the final extension of 5 min. Amplified products were separated in 1.4% agarose gel, stained with ethidium bromide and visualized under ultraviolet (UV) light. The image of gel was taken by multiimager TM 3400 (Alpha Innotech Co.).
Data analysis: RAPD profiles were scored for each individual as discrete characters (presence or absence of amplified products) across all individuals from all populations and for each primer used. Relationships among and between populations were evaluated via the unweighted pair-group method UPGMA (Sneath and Sokal, 1973) and all analysis was performed by using NTSYS (Rohlf et al., 1990). Jaccard’s coefficient was calculated by using FREETREE program, a common estimator of genetic identity and was calculated as follows:
where, NAB is the number of bands shared by samples, NA represents fragments in sample B. Similarity matrices based on these indices were calculated. Similarity matrices were utilized to construct the UPGMA (Unweighted pair group method to construct arithmetic average) dendrogram. Statistical stability of the branches in the cluster was estimated by bootstrap analysis with 1,000 replicates, using the winboot software program (Yap and Nelson, 1996).
RESULTS
Altitudinal variation: Altitudinal variation was found in the genotypes of P. appendiculatum. Genotypes M3, M4 and M5 and M7, M8 and M9 were collected from 4000 and 6000 ft, respectively. Similarly, genotypes M1 (7000ft), M6 (6800 ft) and M2 (6000 ft) showed variation related to their altitude. Thus, these genotypes clearly separated on the basis of their collection site (Fig. 2a) and by band frequency (Fig. 2b) obtained by amplified bands.
RAPD analysis: Genomic DNA amplification of 9 accessions of P. appendiculatum were carried out by using 30 random primers out of which 27 primers yielded 875 reproducible fragments and rest of the 3 primers were not given the amplification (Fig. 3). All the chosen primers amplified across the 9 accessions, with the number of amplified fragments ranging from eighteen (OPA-13) to fourty (OPD-08 and OPAP-03) which varied from 300 to 3500 bp. Out of the 875 amplified bands, 863 were polymorphic, with an average of 29 polymorphic fragments per primer and rest of the 12 fragments were monomorphic (Fig. 3). Percentage of polymorphism ranged from 93.5% (OPD-07) to a maximum 100% with an average of 98.5% polymorphism (Table 3).
The Polymorphism Information Content (PIC) obtained by random amplified bands were obtained with an average of 0.228, ranged from 0.000 to 0.62. Primers, OPA-04, OPA-07, OPA-15, OPA-16, OPA-17, OPA-19 and OPD-10 gave highest PIC values (Table 3).
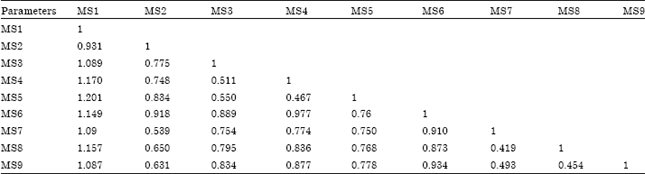
Cluster analysis: Dendrogram based on UPGMA cluster analysis all the 9 accessions of P. appendiculatum were clearly separated according to their altitudinal sites. Genotypes MS1 and MS6 formed separate OUTs from other genotypes according to their high altitude (Fig. 4). However, MS1 and MS6 genotypes appeared to be closer to each other with similarity coefficient of 1.000 and 1.149, respectively (Table 4). Group I consist of genotypes MS3, MS4 and MS5, in which MS3 appeared to closer to M4 with similarity coefficient of 0.511 and genotype MS5 make separate out group with less difference similarity coefficient of 0.550. Similarly, genotype MS2 formed separate out group but appeared close to Group I. In the same manner, Group II consist MS7, MS8 and MS9 genotypes out of which MS7 and MD8 appeared close to each other with similarity coefficient of 0.419 and MS9 formed a separate out group but close to MS7 with similarity coefficient of 0.493. Therefore, all the genotypes are separated in relation to their collection site and altitude.
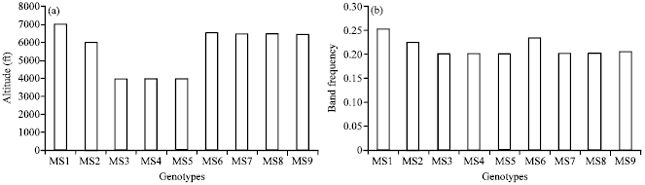 | |
| Fig. 2(a-b): | Graph representing the (a) Altitudinal variation and (b) Band frequency obtained by amplified fragments |
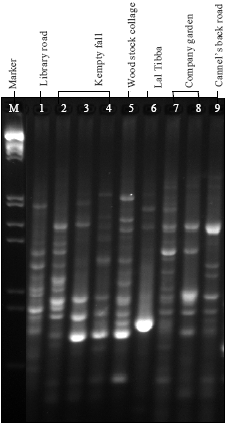 | |
| Fig. 3: | RAPD profile of P. appendiculatum collected from different altitude of Mussoorie. Lane M: EcoRI and Hind III double digested marker. Lane 1: P. appendicualtum collected at 7000 ft; Lane 2: at 6000ft ; Lane 3-5: at 4000ft; Lane 6: at 6800ft; Lane 7-9: at 6400 ft |
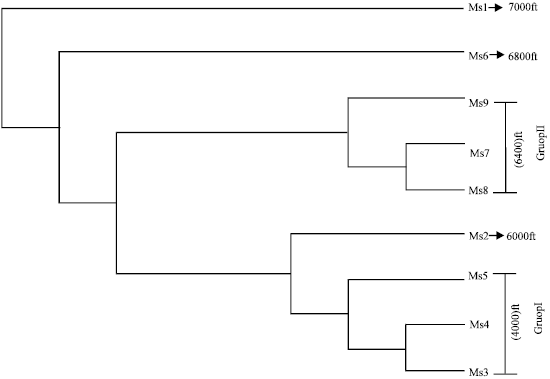 | |
| Fig. 4: | UPGMA based Dendrogram showing the altitudinal variation and genetic relation ship between nine accessions of P. appendiculatum collected from various altitude of Mussoorie |
| Table 3: | Sequences, bands, fingerprints and calculated parameters for the 30 RAPD primer used in Plagiochasma appendiculatum |
 | |
| Table 4: | Jaccard's similarity coefficient matrix followed by UPGMA analysis of P. appendiculatum based on RAPD markers |
 | |
DISCUSSION
The RAPD marker is simple and reproducible technique that allowing comparison of genetic variation between wide range of bryophytes (Boisselier-Dubayle and Bischler, 1989; Bischler-Causse and Boisselier-Dubayle, 1991). Bopp and Capesius (1996) reported that RAPD is used for identification and gene sequencing of bryophytes. However, other methods, such as isozymes and microsatellite have been reported to detect colonization and dispersal in many bryophytes. (Cronberg and Natcheva, 2002; Fang et al., 1997; Stenoien and Sastad, 1999; Rumsey et al., 2001).
The results obtained from the RAPD analysis indicated significant altitudinal variation among the P. appendiculatum, as collected from their different growing site (Fig. 3). UPGMA based dendrogram obtained by RAPD analysis indicated that the genotypes of Group I viz., MS3, MS4 and MS5 comes closer to each other because of short geographical distance and different altitudinal range. On the other hand, similar result has been found within genotypes of Group II i. e., MS7, MS8, MS9. Similarly, genotype MS1 and MS6 are separated according to their higher altitude i.e., 7000 and 6800 ft, respectively (Fig. 4). Soni et al. (2009) deescribed the morphological and genetic variation within and between the genotypes of P. appendiculatum due to growing at different altitude and habitat. These variation is due to the gene flow within closely and far located genotypes. Low gene flow may be one of the major cause of altitudinal variation in P. appendiculatum because high gene rate flow within closly growing individuals occurs by environmental factors indicating not so much vatiation. It means that gene flow takes place from higher to lower altitude and it has been reported that water is essential for the sexual reproduction by mean of dispersal of spores (Wyatt and Anderson, 1984). Restricted gene flow can generally promote local adaptation and genetic divergence between different microhabitats (Via and Lande, 1985). The magnitude of gene flow between the habitats in the current study is difficult to assess. Gamete dispersal distances are considered highly restricted in bryophytes (Wyatt and Anderson, 1984). Spore dispersal distances are probably orders of magnitude higher than gamete dispersal distances.
There is an important role of substrate in bryophyte species diversity and composition has been well established (Pharo and Beattie, 2002), but little is known about the effects of microhabitat and altitude in bryophytes (Romero et al., 2006). Andrew et al. (2003) did not find an overall pattern of bryophyte diversity on different mountains in Tasmania and New Zealand. They considered that altitudinal gradient may control community structure and diversity but suggested that factors operating at smaller scales (moisture, microhabitats) should be studied to understand the underlying mechanisms.
Water plays important role in the spreading of bryophyte propagules from higher to lower altitude along short drainage channels indicating water dispersal where at mixing of geographically divergent populations divided by numerous kilometers and snow confirmed that wind also is imperative for long-range dispersal (Skotnicki et al., 1999). Gene flow in bryophyte is caused during asexual reproduction by dispersal of sperm and spores. All the views of sperm dispersal in mosses and liverworts have reached to the same conclusion: sperm dispersal is very short (Wyatt and Anderson, 1984). Even in large species with splash cups, only rarely do sperms get dispersal more than 50 cm in species without splash cups, fertilization typically occurs within a radius of 10 cm. In the same manner P. appendiculatum growing on short distance having high gene flow so less amount of genetic variation that is why genotypes collected from same altitude comes closer to each other. It is therefore concluded that the RAPD markers may be beneficial for the revealing of altitudinal variation among the genotypes of P. appendiculatum collected from various environmental condition.
ACKNOWLEDGMENTS
The authors are grateful to Director, National Botanical Research Institute, Lucknow India for encouragement and providing facilities and to Department of Biotechnology (DBT), New Delhi for financial support.
REFERENCES
- Andrew, N.R., L. Rodgerson and M. Dunlop, 2003. Variation in invertebrate-bryophyte community structure at different spatial scales along altitudinal gradients. J. Biogeogr., 30: 731-746.
Direct Link - Bischler-Causse, H. and M.C. Boisselier-Dubayle, 1991. Lectotypification of Marchantia polymorpha L. J. Bryol., 16: 361-365.
Direct Link - Boisselier-Dubayle, M.C. and H. Bischler, 1994. A combination of molecular and morphological characters for delimitation of taxa in European Porella. Bryologist, 18: 1-11.
Direct Link - Boisselier-Dubayle, M.C., M.F. Jubier, B. Lejeune and H. Bischler, 1995. Genetic variability in three subspecies of Marchantia polymorpha (Hepaticae): : Isozymes, RFLP and RAPD markers. Taxon, 44: 363-376.
Direct Link - Bopp, M. and I. Capesius, 1996. New aspects of bryophyte taxonomy provided by a molecular approach. Botanica Acta, 109: 368-372.
Direct Link - Cronberg, N. and R. Natcheva, 2002. Hybridization between the peat mosses, Sphagnum capillifolium and S. quinquefarium (Sphagnaceae, Bryophyta) as inferred by morphological characters and isozyme markers. Plant Syst. Evol., 234: 53-70.
CrossRef - Frahm, J.P. and R. Ohlemuller, 2001. Ecology of bryophytes along altitudinal and latitudinal gradients in New Zealand. Studies in austral temperate rain forest bryophytes 15. Trop. Bryol., 20: 117-137.
Direct Link - Frahm, J.P., 1990. The altitudinal zonation of bryophytes on Mt. Kinabalu. Nova Hedwigia, 51: 133-149.
Direct Link - Rumsey, F.J., J.C. Vogel and S.J. Russell, 2001. A study of genetic variation in the threatened hepatic Petalophyllum ralfsii (Wils.) Nees. and Gottsche (Fossombroniaceae). Conserv. Genet., 2: 271-277.
CrossRef - Freitas, H. and A. Brehm, 2001. Genetic diversity of the macaronesian leafy liverwort Porella canariensis inferred from RAPD markers. J. Heredity, 92: 339-345.
Direct Link - Ghate, S. and S.B. Chaphekar, 2000. Plagiochasma appendiculatum as a biotest for water quality assessment. Environ. Pollut., 108: 173-181.
CrossRef - Gradstein, S.R. and J. Frahm, 1987. Die floristische Hohengliederung der Moose entlang des BRYOTROP-Transektes in NO-Peru. Beihefte zur. Nova. Hedwigia, 88: 105-113.
Direct Link - Kessler, M., 2000. Altitudinal zonation of Andean cryptogam communities. J. Biogeogr., 27: 275-282.
CrossRef - Kumar, K., K.K. Singh, A.K. Asthana and V. Nath, 2000. Ethnotherapeutics of bryophyte Plagiochasma appendiculatum among the Gaddi tribes of Kangra Valley, Himachal Pradesh, India. Pharm. Biol., 38: 353-356.
CrossRef - Mishler, B.D., 2001. The biology of bryophytes-bryophytes aren’t just small tracheophytes. Am. J. Bot., 88: 2129-2131.
Direct Link - Pharo, E.J. and A.J. Beattie, 2002. The association between substrate variability and bryophyte and lichen diversity in eastern Australian forests. Bryologist, 105: 11-26.
CrossRefDirect Link - Pharo, E.J., A.J. Beattie and D. Binns, 1999. Vascular plants diversity as a surrogate for bryophyte and lichen diversity. Conserv. Biol., 13: 282-292.
Direct Link - Rohlf, E.J., W.S. Chang, R.R. Sokal and J. Kim, 1990. Accuracy of estimated phylogenies: Effects of tree topology and evolutionary model. Evolution, 44: 1671-1684.
Direct Link - Romero, C., F.E. Putz and K. Kitajima, 2006. Ecophysiology in relation to exposure of pendant epiphytic bryophytes in the canopy of a tropical montane oak forest. Biotropica, 38: 35-41.
CrossRef - Selkirk, P.M., M. Skotnicki, K.D. Adam, M.B. Connett, T. Dale, T.W. Joe and J. Armstrong, 1997. Genetic variation in Antarctic populations of the moss Sarconeurum glaciale. Polar Biol., 18: 344-350.
CrossRefDirect Link - Singh, M., R. Govindarajan, V. Nath, A.K.S. Rawat and S. Mehrotra, 2006. Antimicrobial, wound healing and antioxidant activity of Plagiochasma appendiculatum Lehm. et Lind. J. Ethnopharmacol., 107: 67-72.
CrossRefDirect Link - Skotnicki, M.L., J.A. Ninham and P.M. Selkirk, 2000. Genetic diversity, mutagenesis and dispersal of Antarctic mosses: A review of progress with molecular studies. Antarctic Sci., 12: 363-373.
CrossRefDirect Link - Skotnicki, M.L., P.M. Selkirk and C. Beard, 1998. RAPD profiling of genetic diversity in two populations of the moss Ceratodon purpureus in Victoria Land, Antarctica. Polar Biol., 19: 172-176.
CrossRefDirect Link - Skotnicki, M.L., P.M. Selkirk and J.A. Ninham, 1999. RAPD analysis of genetic variation and dispersal of the moss Bryum argenteum in Ross Island and Victoria land, Antarctica. Polar Biol., 21: 417-422.
CrossRefDirect Link - Scotnicki, M.L., P.M. Selkirk, P. Broady, K.D. Adam and J.A. Ninham, 2001. Dispersal of the moss Campylopus pyriformis on geothermal ground near the submits of Mount Erebus and Mount Melbourne, Victoria Land, Antarctica. Antarctic Sci., 13: 280-285.
Direct Link - Stenoien, H.K. and S.M. Sastad, 1999. Genetic structure in three haploid peat mosses (Sphagnum). Heredity, 82: 391-400.
CrossRef - Via, S. and R. Lande, 1985. Genotype-environment interaction and the evolution of phenotypic plasticity. Evolution, 39: 505-522.
CrossRefDirect Link - Wigginton, M.J., 2002. Checklist and distribution of the liverwort and hornworts of sub-Saharan Africa, including the East African Islands. Trop. Bryol. Res. Rep., 3: 1-88.
Direct Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Soni, A., A. Kumar, V. Nath and A. Niveden, 2009. Genetic diversity of indian liverwort Plagiochasma appendiculatum revealed by RAPD marker. Res. J. Botany, 4: 89-100.
CrossRefDirect Link - Soni, A. and A. Kumar 2009. Protocol for improved extraction and PCR amplification of genomic DNA from liverwort, Plagiochasma appendiculatum. Indian J. Exp. Biol., 47: 921-924.
PubMedDirect Link