
Abdulhakeem A. Al-Majed
Department of Pharmacology, College of Pharmacy, King Saud University, P.O. Box 2457, Riyadh-11451, Saudi Arabia
Journal of Medical Sciences
Year: 2007 | Volume: 7 | Issue: 2 | Page No.: 276-283







ABSTRACT
Valerian (a natural psychotropic) has world-wide folkloric significance as anti-anxiety and a tranquilizing herbal drug. In view of its immense use and a paucity of literature on its toxicity, valerian was evaluated for its effects on spermatogenesis, genotoxicity, reproduction and biochemical changes in male Swiss albino mice. The mice were orally treated with different doses (125, 250 and 500 mg/kg/day) of the aqueous suspension of valerian for 90 days. The following parameters were studied: (1) reproductive organ weight (2) motility and content of sperms (3) spermatozoa morphology (4) cytology of the testis chromosomes (5) study on reproduction (6) biochemical study on proteins, nucleic acids, malonaldehyde (MDA) and non-protein sulfhydryl (NP-SH). The treatment induced significant increase in the weight of caudae epididymis and seminal vesicles and sperm count. There was no effect on sperm motility, morphology of spermatozoa, rate of pregnancy and pre- and post- implantation losses. However, the treatment was found to increase the frequency of chromosomal aberrations. The study on biochemical parameters showed depletion of nucleic acids, NP-SH and increase of MDA, which elucidated the role of free radical species in the induction of chromosomal aberrations. The influence of biochemical changes on induction of chromosomal aberrations and the lack of any effect on spermatogenic and reproductive function is discussed.
PDF Abstract XML References
How to cite this article
Abdulhakeem A. Al-Majed, 2007. Effect of Valerian on Spermatogenic, Genotoxic,Reproductive and Biochemical Changes in Sex Cells after Chronic Treatment in Male Swiss Albino Mice. Journal of Medical Sciences, 7: 276-283.
DOI: 10.3923/jms.2007.276.283
URL: https://scialert.net/abstract/?doi=jms.2007.276.283
DOI: 10.3923/jms.2007.276.283
URL: https://scialert.net/abstract/?doi=jms.2007.276.283
INTRODUCTION
Valerian (Valerianaceae) an herbal product is reported to possess sedative, anti-anxiety and tranquilizing properties (Boniel and Dannon, 2001; Cas, 2004) and is used for the treatment of anxiety, insomnia and restlessness (Birgit et al., 2005). It has many adverse effects, such as, mydriasis, nephrotoxicity, tremors, headache, abdominal pain, congestion in chest and nephrotoxicity (Boniel and Dannon, 2001). Studies on hematology revealed valerian to increase bleeding tendency (Ang-Lee et al., 2001; Hodges and Kam, 2002), cause spontaneous hemorrhage (Boniel and Dannon, 2001) and produce coagulation disorders (Cheng et al., 2002). Higher doses of valerian extract are found to induce DNA damage in human endothelial cell line (ECV304) (Hui-Lian et al., 2003).
The therapeutic activity and toxicity of valerian is attributed to different constituents present in it. Valepotriates (valiracyl, valtrate/isovaltrate and dihydrovaltrate) are considered to be the main tranquilizing constituents of valerian. They are found to possess a strong alkylating activity against the nucleophilic agent 4-(p-nitrobenzyl)-pyridine (Von der Hude et al., 1985). Dunaev et al. (1987) found valiracyl to exert neurotropic effect. This effect is known to inhibit the levels of GABA and decrease the bioenergetic processes in brain. Aglycones (prinsepiol and chlorogenic acid), valepotriates (valtrate/isovaltrate), dihydrovaltrate, baldrinal and homobaldrinal are reported to be mutagenic in Salmonella typhimurium (TA 100) and E. coli WP2 and WP2 (Von der Hude et al. (1985, 1986) and cytotoxic in cultured hepatoma cells (Keochanthala-Bounthanh et al., 1990).
Although a large number of papers are published on the adverse effects of valerian and its constituents, little is known about their effect on the spermatogenesis and reproductive function, except the reports on the effect of valepotriates on fertility index in female rats (Tufik et al., 1994). The present study was undertaken in view of a paucity of literature on the effect of valerian on spermatogenesis, genotoxicity, reproductive function and biochemical changes in sex cells. The rationale of this study is based on (1) the proven CNS depressant activity of valerian (Leuschner et al., 1993) and the reported sexual dysfunction caused by CNS depressants and sedating drugs (Petrie, 1980; Aldridge, 1982; Fabbri et al., 2003) (2) the observed vaso-relaxant activity of aglycone constituents (Piccinelli et al., 2004) and the role of blood supply to the penis in erectile function (Fabbri et al., 2003).
MATERIALS AND METHODS
The present study on genetic and biochemical effects of Valerian was conducted in the Department of Pharmacology, College of Pharmacy, King Saud University. The experimental part was undertaken during the period Nov 05 to July 06.
Test herbal product: Valerian was used as the test herbal product in the present study. It is marketed by Nature’s way product (USA) in form of capsules. Each capsule contained 800 mg (valerian root) and 220 mg, valerian dried extract.
Animal stocks: Male Swiss albino mice (SWR) aged 6-8 weeks and weighing 25-28 g were obtained from the Experimental Animal Care Center, King Saud University, Riyadh, Saudi Arabia. The animals were fed on Purina chow diet and water ad libitum and were maintained under standard conditions of humidity, temperature and light (12 h, light/12 dark cycle). The conduct of experiments and the procedure of sacrifice (using ether) were approved by the Ethics Committee of the Experimental Animal Care Society, College of Pharmacy, King Saud University, Riyadh, Saudi Arabia.
Dose, route and duration of treatment: The doses of valerian selected were 125, 250 and 500 mg/kg/day corresponding to 1/64, 1/32 and 1/16, respectively of the evaluated maximum tolerated dose (8.0 g kg-1), (Chan et al., 1986). Aqueous suspension of valerian was administered by gastric intubation (oral) daily for a period of 90 days.
Experimental groups: The experimental groups of mice consisted of the following: group 1, control (tap water); group 2, valerian (125 mg/kg/day); group 3, valerian (250 mg/kg/day); group 4, valerian (500 mg/kg/day). In each group, a total of 25 male mice were used. In studies on evaluation of reproductive performance 30 female mice were used in each group. The allotment of mice for different experiments was as follows: (I) studies on spermatogenic function (5 mice); (ii) biochemical evaluation (5 mice); (iii) cytological procedures (5 mice) and (iv) evaluation of reproductive performance (10 male mice and 30 female mice).
Evaluation of spermatogenic function: The mice were sacrificed after the last day of sub-chronic treatment (Wyrobek and Bruce, 1975; Al-Shabanah, 1997) and weighed for the essential reproductive organs, such as, testis, caudae epididymis, seminal vesicles and prostate glands. The spermatozoa were obtained by making small cuts in caudae epididymis and vas deferens placed in 1 mL of modified Krebs Ringer-bicarbonate buffer (pH 7.4). After 10 min, the epididymis and vas deferens tubules were removed and the resultant sperm suspension evaluated for sperm content, percent motility. The percent motility of sperms was determined by their progressive and non-progressive movements observed under a compound microscope (Anderson et al., 1983). The sperm count was determined under a Neubauer haemocytometer (Wyrobek et al., 1983). The sperm suspension obtained was stained with 0.05% of eosin-Y, smears were made on slides, air-dried and made permanent. The spermatozoal morphology was examined by bright-field microscopy with an oil immersion lens. The different spermatozoal abnormalities (amorphous, banana shaped, swollen achrosome, triangular head, macrocephali and rotated head) screened were those found in all the slides (Wyrobek and Bruce, 1975; Al-Shabanah, 1997).
Cytogenetic analysis for meiotic chromosomes: In the analysis of the chromosomal aberrations, the mice were sacrificed after the last day of the sub-chronic treatment (Schleismacher, 1970; Al-Shabanah, 1997). The testis was removed in an isotonic sodium citrate solution. After peeling out the tunica albugenia, the seminiferous tubules were teased to form a cell suspension. The cell suspension was centrifuged and the pellet re-suspended in the hypotonic citrate solution. After the second centrifugation the supernatant was discarded and the pellet suspended in fixative (methanol and acetic acid, 3:1). The chromosomal preparations were made by the air drying technique (Evans et al., 1964; Al-Shabanah, 1997). The coded slides were stained in Giemsa solution (10%) and screened for the chromosomal aberrations including aneuploids, autosomal univalents, sex-univalents, polyploids and translocations.
Reproductive performance: The dominant lethal assay was carried to study the reproductive performance of male mice (Green et al., 1985; Al-Shabanah, 1997). After treatment, each male mouse in the treated and control groups was caged with three females for mating. After 1 week, females were replaced by another three fresh virgin female mice per male, which were also being allowed to stay with the male for one week. The female mice were necropsied 13 days following the mid-week of their caging and presumptive mating and the number of pregnant mice was recorded to determine percent fertility (Al-Shabanah, 1997). The pre-implantation loss was calculated by comparing the number of implantations per pregnant female in the treated and control groups.
The post-implantation loss was determined by the number of dead implantations per pregnant female as a measure of dominant lethality (Al-Shabanah, 1997; Agrawal et al., 1997).
Estimation of total proteins: Total proteins were estimated by the modified Lowry method of Schacterle and Pollack (1973). Bovine serum albumin was used as standard.
Determination of nucleic acids: The method described by Bregman (1983) was used to determine the levels of nucleic acids. Testes were homogenized and the homogenate was suspended in ice-cold trichloroacetic acid (TCA). After centrifugation, the pellet was extracted with ethanol. DNA was determined by treating the nucleic acid extract with diphenylamine reagent and reading the intensity of blue color at 600 nm. For quantification of RNA, the nucleic acid extract was treated with orcinol and the green colour was read at 660 nm. Standard curves were used to determine the amounts of nucleic acids present.
Malondialdehyde estimation: The method described by Ohkawa et al. (1979) was used. Malondialdehyde (MDA) was measured as an indicator of lipid peroxidation. Testes were homogenized in KCl solution and incubated with thiobarbituric acid. After centrifugation the pink clear layer was read at 532 nm. Malondialdehyde bis (dimethyl acetal) was used as an external standard.
Determination of nonprotein sulfhydryl groups (NP-SH): The estimation of NP-SH levels in different organs was undertaken according to the method of Sedlak and Lindsay (1968). The testes were homogenized in ice cold 0.02 M ethyleneodiaminetetraacetic acid disodium. The homogenate was treated with 50% w/v trichloroacetic acid and centrifuged. Supernatant fractions were mixed with tris buffer, 5-5’-dithiobis-(2 nitrobenzoid acid) (DTNB) was added. After shaking the contents, its absorbance was determined at 412 nm within 5 min of the addition of DTNB against reagent blank with no homogenate.
Statistical analysis: The different studies undertaken were statistically analyzed by Analysis of variance. Some parameters in studies on reproductive performance were analyzed using a Chi-square test.
RESULTS
The sub-chronic treatment with valerian showed a significant increase in the weight of caudae epididymis and seminal vesicles (p<0.05) at the high dose (500 mg/kg body weight/day) as compared to the values obtained in the control (Fig. 1).
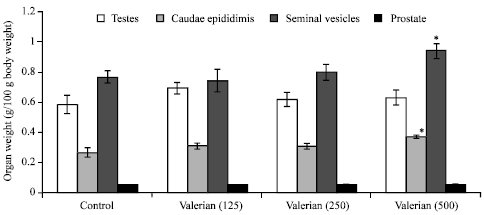 | |
| Fig. 1: | Effect of valerian (mg/kg body weight/day) on reproductive organ weight in male Swiss albino mice, Five mice were used in each group, *p<0.05 (one-way ANOVA and Post-hoc Tukey-Kramer multiple comparison test was done) |
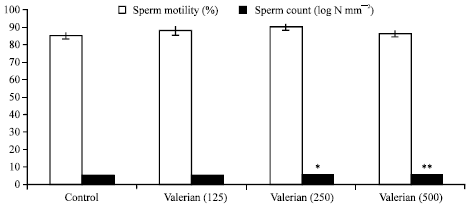 | |
| Fig. 2: | Effect of valerian (mg/kg body weight/day) on sperm motility and count in Swiss albino mice after sub-chronic treatment, Five mice were used in each group, *p<0.05, **p<0.01 (one-way ANOVA and Post-hoc Tukey-Kramer multiple comparison test was done) |
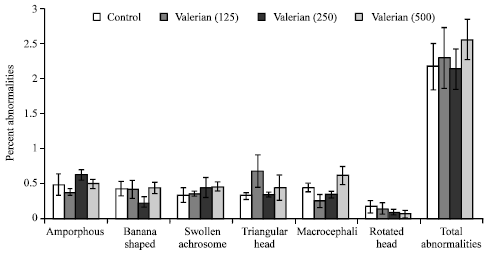 | |
| Fig. 3: | Effect of valerian (mg/kg body weight/day) on epididymal spermatozoa in Swiss albino mice after sub-chronic treatment, Five mice were used in each group, p>0.05 (one-way ANOVA and Post-hoc Tukey-Kramer multiple comparison test was done) |
The treatment also caused significant increase in the sperm count at 250 (p<0.05) and 500 (p<0.01) mg/kg body weight/day, while there were no such changes observed in the percent sperm motility (Fig. 2).
The prolonged treatment with valerian failed to significantly affect any of the morphological abnormalities in epididymal spermatozoa (amorphous, banana shaped, swollen achromosome, triangular head, macrocephali and rotated head) of mice (Fig. 3).
| Table 1: | Effect of valerian on the induction of dominant lethal mutations after sub-chronic treatment in male Swiss albino mice |
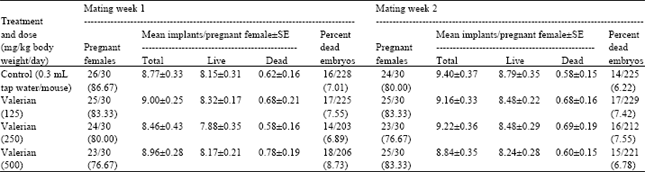 | |
| Figures between parentheses denote percent, A total of 10 male and 30 females were used in each group, p>0.05 (One way ANOVA and Post hoc Tukey-Kramer multiple comparison test), p>0.05 (Chi square test) | |
| Table 2: | Effect of valerian on proteins, nucleic acids, malondialdehyde and glutathione (NP-SH) contents in testicular tissue of male Swiss albino mice after Sub-chronic treatment |
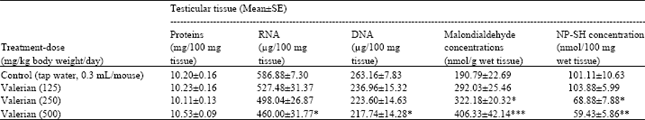 | |
| Five mice were used in each group, *p<0.05; **p<0.01; ***p<0.001 (One way ANOVA and Post hoc Tukey-Kramer multiple comparison test was done) | |
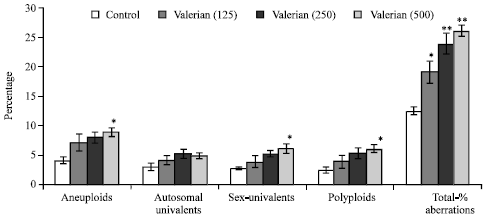 | |
| Fig. 4: | Effect of valerian (mg/kg body weight/day) on testis chromosomes in Swiss albino mice after sub-chronic treatment, Five mice were used in each group, *p<0.05, **p<0.01 (one-way ANOVA and Post-hoc Tukey-Kramer multiple comparison test was done) |
Valerian treatment for long term caused significant (p<0.05) changes in the frequency of aneuploids, sex-univalents, polyploids at the high dose (500 mg/kg/day). The total percent aberrations were also significantly increased at all the different doses 125 (p<0.05) and 250 and 500 (p<0.01) of valerian (Fig. 4).
The sub-chronic treatment with valerian failed to decrease the rate of pregnancy in any of the mating weeks. The treatment did not cause any change in the pre- and post- implantation loss or the percent dead embryos (Table 1).
Prolonged treatment with valerian induced significant (p<0.05) increase in the concentrations of nucleic acids (RNA and DNA) at the high dose (500 mg/kg body weight/day) as compared to the values obtained in the control. The concentrations of MDA were significantly (p<0.05) increased at the medium dose (250 mg/kg body weight/day) and (p<0.001) at the high dose (500 mg/kg body weight/day). The levels of NP-SH were significantly decreased at the higher doses 250 (p<0.05) and 500 (p<0.01) mg/kg body weight/day (Table 2).
DISCUSSION
The sub-chronic treatment with valerian significantly increased the weight of caudae epididymis and seminal vesicles. The frequency of sperm count was also increased. However, the percent motility, morphology of spermatozoa, male fertility, pre- and post- implantation losses were not affected. There is a paucity of related literature on valerian, hence a comparison is difficult. However, the data on sperm count support literature reports (Petrie, 1980) which suggest CNS depressants and sedatives (like valerian) to cure oligospermia by improving sperm count. The lack of any effect on male fertility, pre- and post- implantation losses are not in corroboration with the reported sexual dysfunction associated with CNS-depressants and sedatives (Aldridge, 1982; Fabbri et al., 2003). The discordance in these results might be due to vaso-relaxant property of aglycones (prinsepiol and chlorogenic acid) present in valerian (Piccinella et al., 2004).
The results on testes chromosomes showed significant increase in different types of aberrations. These data are confirmed by the significant depletion of both RNA and DNA, observed in the present study. There is no parallel study on genotoxicity of valerian, except the induction of DNA damage, observed in human endothelial cell line (ECV302) (Hui-Lian et al., 2003). However, a comparison between these reports is difficult, in view of the differences in the sensitivity of normal and cancerous cells. The observed effect on testicular chromosomes and biochemical changes in the testis might be due to the presence of some tranquilizing constituents (valtrate/isovaltrate and dihydrovaltrate) in valerian and their metabolites (baldrinal and homobaldrinal) (Von der Hude et al., 1986). Valtrate/isovaltrate and dihydrovaltrate have been found to be mutagenic in Salmonella typhimurium after metabolic activation in presence of S9-mix, whereas, baldrinal and homobaldrinal showed genotoxicity with and without metabolic activation (Von der Hude et al., 1986). The depletion of testicular nucleic acids might be ascribed to the non-volatile monoterpenes (valepotriates) present in valerian. Earlier studies (Keochanthala-Bouthanh et al., 1990) demonstrated these terpenes to be cytotoxic in cultured hepatoma cells. The exact mode of action of the observed effect on testis chromosomes and the depletion of nucleic acids is not known; however, it might be related with the generation of free radical species, as revealed by the increase and depletion of MDA and NP-SH, respectively in the testicular cells, which clearly demonstrated the oxidant potentials of valerian in testicular cells. Previous reports suggested that the deficiency of endogenous antioxidants interfere with meiosis and cause chromosomal anomalies (Watanabe and Endo, 1991; Al-Shabanah, 1997). Nevertheless, these data contradict the literature reports on a large number of constituents (terpenes, lignans, flavonoids [kaempferol, quercetin], isoflavonoids, caretonoids) of valerian that are found to be antioxidants and known to protect against genotoxicity and/or carcinogenicity (Harper et al., 1999; Osswald et al., 2000; Malva et al., 2004). The discordance in these results might be due to the redox state of the cells and the pre-loading of valerian much before the exposure of the tissues to oxidant injury. The difference in duration and quantity of exposures might have determined the proportion of antioxidant/oxidant ratio and the related outcome. Another possibility may be that the pharmacokinetic and pharmaco-dynamic interactions between the multiple constituents and their metabolites might also be responsible for the toxic potentials of valerian.
Taken together, this study has revealed that valerian has no effect on the spermatogenic cycle, spermatozoa morphology and the reproduction, but there was a significant increase in the frequency of chromosomal aberrations and concentration of MDA and decrease in the levels of nucleic acids and NP-SH in the testes. The discordance between these results might be related to the role of the excurrent duct system in eliminating morphologically abnormal spermatozoa during transit from testes to vas deferens and caudae epididymis. Previous studies (Perez-Sanchez et al., 1997; Chenoweth et al., 2000) have shown that the transit of spermatozoa through the extragonadal passage produces and/or eliminate morphologically abnormal spermatozoa by the excurrent duct system. The mechanism of these changes is yet a dilemma.
The exact mechanism of action of valerian-induced genotoxicity and the biochemical changes is not known. However, it appears to be to be related with the influence of valerian to suppress the cytochrome P450 enzymes (Lefebvre et al., 2004) and cause the related accumulation of oxidative damage (Sapone et al., 2003; Wauthier et al., 2004; Imaoka et al., 2004). These changes might have increased the susceptibility of the germ cells (Montero et al., 2003) to the observed genetic and biochemical changes. The observed genotoxicity and the related biochemical changes might be attributed to either one constituent or it may be the cumulative effect of the different oxidant constituents (valtrate/isovaltrate, dihydrovaltrate, baldrinal and homobaldrinal) of valerian (Vonder Hude et al., 1985; Hui-Lian et al., 2003; Keochanthala-Bounthanh et al., 1990).
It is concluded that valerian cause biochemical changes which might be responsible for chromosomal aberrations in the testes. However, lack of any effect on sperm morphology, pre- and post implants show that the chromosomal anomalies may not have been carried to the mature spermatozoa available for fertilization. It is expected that the morphologically abnormal spermatozoa (if formed) might have been eliminated by the excurrent duct system in their transit through extra gonadal passage from testis to vas deferens and caudae epididymis. Further experiments on molecular mechanisms are in progress.
ACKNOWLEDGMENT
The author is thankful to King Abdulaziz City for Science and Technology, Riyadh, Kingdom of Saudi Arabia for sanction of the grant to conduct research on herbal drugs (KACST RESEARCH PROJECT No. AR-21-41).
REFERENCES
- Ang-Lee, M.K., J. Moss and C.S. Yuan, 2001. Herbal medicines and perioperative care. J. Am. Med. Assoc., 286: 208-216.
CrossRef - Cheng, B., C.T. Hung and W. Chiu, 2002. Herbal medicine and anaesthesia. Hong Kong Med. J., 8: 123-130.
Direct Link - Chenoweth, P.J., C.C. Jr. Chase, C.A. Risco and R.E. Larsen, 2000. Characterization of gossypol-induced sperm abnormalities in bulls. Theriogenology, 93: 1193-1203.
Direct Link - Evans, E.P., G. Breckon and C.E. Ford, 1964. An air-drying method for meiotic preparations from mammalian testes. Cytogenetics, 3: 289-294.
PubMedDirect Link - Fabbri, A. and M. Caprio, 2003. Pathology of erection. J. Endocrinol. Invest., 26: 87-90.
Direct Link - Harper, A., D.J. Kerr, A. Gescher and J.K. Chipman, 1999. Antioxidant effects of isoflavonoids and lignans and protection against DNA oxidation. Free Rad. Res., 31: 149-160.
Direct Link - Hui-Lian, W., Z. Dong-Fang, L. Zhao-Feng, L. Yang, L. Qian-Rong and W. Yu-Zhen, 2003. In vitro study on the genotoxicity of dichloromethane extrcts of valerian (DEV) in human endothelial ECV304 cells and the effect of vitamins E and C in attenuating the DEV-induced DNA damages. Toxicol. Applied Pharmacol., 188: 36-41.
Direct Link - Imaoka, S., M. Osada, Y. Minamiyama, T. Yukimura, S. Toyokuni, S. Takemura, T. Hiroi and Y. Funae, 2004. Role of Phenobarbital-inducible cytochrome P450s as a source of active oxygen species in DNA-oxidation. Cancer Lett., 203: 117-125.
Direct Link - Lefebvre, T., B.C. Foster, C.E. Drouin, A. Krantis, J.F. Livesey and S.A. Jordan, 2004. In vitro activity of commercial valerian root extracts against human cytochrome P450 3A4. J. Pharm. Pharmaceut. Sci., 7: 265-273.
Direct Link - Montero, R., L. Serrano, V.M. Davita and A. Ito, 2003. Plancarte: Infection of rats with Taenia taeniformis metacestodes increases hepatic CYP450, induces the activity of CYP1A1, CYP2B1 and COH isoforms and increases the genotoxicity of the procarcinogens benzo[a]pyrene, cyclophosphamide and aflatoxin B(1). Mutagenesis, 18: 211-216.
Direct Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Osswald, K., T.W. Becker, M. Grimm, G. Jahreis and B.L. Pool-Zobel, 2000. Inter- and intra-individual variation of faecal water genotoxicity in human colon cells. Mutat. Res., 472: 59-70.
Direct Link - Perez-Sanchez, F., L. Tablado and C. Soler, 1997. Sperm morphological abnormalities appearing in the male rabbit reproductive tract. Theriogenology, 47: 893-901.
Direct Link - Piccinelli, A.L., S. Arana, A. Caceres, R. di Villa Bianca, R. Sorrentino and L. Rastrelli, 2004. New lignans from the roots of Valeriana prionophylla with antioxidative and vasorelaxant activities. J. Natural Prod., 67: 1135-1140.
Direct Link - Sapone, A., A. Affatato, D. Canistro, M. Broccoli and S. Trespidi et al., 2003. Induction and suppression of cytochrome P450 isoenzymes and generation of oxygen radicals by procymidone in liver, kidney and lung of CD1 mice. Mutat. Res., 527: 67-80.
Direct Link - Schacterle, G.R. and R.L. Pollack, 1973. A simplified method for quantitative assay for small amount of proteins in biological materials. Anal. Biochem., 51: 654-655.
PubMed - Sedlak, J. and R.H. Lindsay, 1968. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman's reagent. Anal. Biochem., 25: 192-205.
CrossRefPubMedDirect Link - Tufik, S., K. Fujita, L. Seabra, L. Mde and L. Lobo, 1994. Effects of a prolonged administration of valepotriates in rats on the mothers and their offspring. J. Ethnopharmacol., 41: 39-44.
Direct Link - Wauthier, V., R.K. Verbeeck and P.B. Calderon, 2004. Age-related changes in the protein and mRNA levels of CYP2E1 and CYP3A isoforms as well as in their hepatic activities in Wistar rats. What role for oxidative stress. Arch. Toxicol., 78: 131-138.
Direct Link - Wyrobek, A.J. and W.R. Bruce, 1975. Chemical induction of sperm abnormalities in mice. Proc. Natl. Acad. Sci., 72: 4425-4429.
CrossRefPubMedDirect Link - Wyrobek, A.J., L.A. Gordon, J.G. Burkhart, M.W. Francis and R.W. Kapp et al., 1983. An evaluation of the mouse sperm morphology test and other sperm tests in nonhuman mammals: A report of the U.S. environmental protection agency Gene-Tox program. Mutat. Res./Rev. Genet. Toxicol., 115: 1-72.
CrossRefPubMedDirect Link