
Ambalavana Ganapathi Pillay
St. Matthews
University School of Medicine,
Safe-Haven, Leeward Three
Post Box, 32330 SMB,
Grand Cayman (BWI), UK
Journal of Medical Sciences
Year: 2003 | Volume: 3 | Issue: 5 | Page No.: 367-394







ABSTRACT
Senescent-decline in the nervous-system functions is very frequently attributed to age-related neurone-loss. The major part of this work is aimed at analysing this behaviour of the neuronal elements during the whole life of a single animal species. On the evidence available during development of different ganglia in the present study, the dark cells are considered as active ones; the light cells are considered as those which have failed to establish proper functional projections, inactive, dying, dead or degenerating ones. Probably it is during the early stages of cell growth (small and medium-sized ones = 11B 20μ size), the peripheral and central processes (of axons) begin to grow from the cell body and attempt to get established in their projection fields. If they fail in this attempt they lose their activity, tend to die and disappear, and change to a light coloured cell on staining. The tiny cells are always found to be dark type expressing their most active period. The very-small type of cells is also usually dark during embryonic development till E18. The light cells have appeared among the very small type of cells also just on the day of hatching that may or may not continue in the adult situation. This probably signifies the possible attempt to eliminate the growing cells since they are no longer needed to replace larger categories of cells which have already well-developed neuronal connections at this stage. On E18, there is tremendously increased number of phagocytic cells while the light cells have greatly reduced in number. The phagocytosis is too fast that the inactive, dying or dead cells (i.e., so-called light cells) are removed immediately as soon as they are formed. The light cell stage becomes clearly observable only when some of the important connections are being actively established. It is assumed from these observations that the time of appearance of light cells might be indirectly related to the onset of establishment of active functional connections of neurones and to the functional importance of the organs which it supplies.
PDF Abstract XML References Citation
How to cite this article
Ambalavana Ganapathi Pillay, 2003. Developmental and Growth Changes in Neuron Differentiation, Dark and Light Neurons, and Age-related Neuronal Death in the Cranial Nerve Ganglia and in the Autonomic Nervous System with Reference to their Functional Significance: A Contribution to the Neurosensory and Motor Control of Living, Habits, Behaviour and Aging Process. Journal of Medical Sciences, 3: 367-394.
DOI: 10.3923/jms.2003.367.394
URL: https://scialert.net/abstract/?doi=jms.2003.367.394
DOI: 10.3923/jms.2003.367.394
URL: https://scialert.net/abstract/?doi=jms.2003.367.394
INTRODUCTION
Senescent-decline in the nervous-system functions is very frequently attributed to age-related neurone loss. Processes and mechanisms involved in neuro-degeneration form part of the structural framework for interpreting the functional consequences of age-related changes in other parameters. The literature concerning age-related neurone loss give confusing and sometimes contradicting data and therefore, remain with controversy (Coleman and Flood 1987, Bondareff 1987, Dlugos and Pentney 1994, Heinensen et al 1994, West 1993). However, age-related neurone-loss represents a structural basis of senescent-decline in nervous system functions. Dark and Light types of neurones, based on staining properties have been documented in many vertebrates (Dixon 1963, Moses et al 1965, Peach 1972, Meyer et al 1973) including primates (Kerr 1967, Carmel and Stein 1969). A few investigators (Peach 1972, Kalina and Wolman 1970, Silbermann and Finkelbrand 1978) have found differences in chemical constituents in these two types of neurones in sensory ganglia of rodents. Similar observations have been reported in mammals (Peach 1972, Cauna and Naik 1963, Kalina and Bubis 1969) and reptiles (Kishida et al 1982).
The significance of Dark and Light neurones in different ganglia in different animal species has been controversial in available literature. Dual embryonic origin (of epidermal placode and of neural crest origin) (Hamburger 1961), as fixation artifacts (Cammermeyr 1962), difference in central and peripheral projections (Preto Parvis 1954, Gobel 1974), different sensory functions (Noden 1980, Spassova 1982), different histogenetic characteristics (Meyer 1973), difference in distribution of cytoplasmic organelles and relative density of cytoplasm (Moses et al 1965, Carmel and Stein 1969, Matsura et al 1969), fluid shift between cells and the surrounding extra-cellular spaces (Moses 1967) have been offered as different hypotheses. From available literature, there is no report of a study in the whole life cycle of any one animal species so as to infer a conclusive significance of this dual cytology of neurones. All these works have been done in adult animals or in certain stages of development or growth.
Therefore, it is thought useful to study this aspect in different ganglia related to different functions, in the whole life cycle of any one animal species during embryonic development through adult, so as to infer the conclusive significance and hypothesis regarding the occurrence of this dual cytology and to see whether their occurrence is related to age-changes. This has been really rewarding to achieve this conclusive information from the results obtained. In this article, the discussion is restricted to some useful points (rather than description) in order to simplify a great deal of repetition (the detailed descriptions in relation to individual ganglia may be found elsewhere : Pillay 1999 a, b, c, d, e; 2000 a, b, c, 2001 a, b, c). The results in the present study can be of great value to correlate and evaluate the functional status of similar ganglia during development and in relevant clinical conditions and ageing in the human.
MATERIALS AND METHODS
The chicks gallus gallus domesticus, Whit leghorn breed were used. Fertilized eggs were incubated at 37.5o C, after every 24 hours, it was considered as Embryonic day 1 (E1), Embryonic day 2 (E2) etc till hatching (H). Embryos / Fetuses till hatching were removed carefully under anesthesia and aseptic conditions, and fixed in 10% formaldehyde solution (HCHO) solution at least for two weeks. Large (older) fetuses were cut transversely into suitable smaller pieces and labelled serially for future orientation. The tissues of older fetuses (i.e., E 15 and onwards till adult) were usually decalcified after fixation. Serial sections of 8 B 10 μ were stained by Cresyl Fast Violet for Nissl granules. Only a few selected stages that showed some remarkable changes i.e., E 6, E 8,E 10, E 13 E 15, E 18, chicks on the day hatching (H) and adult are included in this investigation. In all three animals in each group, with a total of twenty-four animals were used.Ganglia of both sides were used for examination. Every section of the ganglion was examined, drawn and the cells were plotted in a diagram with the help of a light microscope having acamera lucida attachment. Different categories of neurons were classified into Dark and Light neurons according to the difference in the intensity of cytoplasmic stain (Pillay, 1999a). Only those cells having a clear nucleus, and a nucleolus were counted and measured with the help of an eyepiece graticule. The following categories of cells were classified.
Tiny (< 5 μ), very small (6B10 μ), small (11B15 μ), medium sized (16B20 μ), big (21B25 μ), very big (26B30 μ), large (31B35 μ), very large (36B40 μ), giant (41B45 μ) (Pillay, 1999a). The categorization of cells on the basis of size with uniform difference of 5 μ was initially maintained just for the sake of convenience. However, this proved to be very useful in that, the behaviour, especially that of the very small cells, is very interesting on the day of hatching (H) (uniformly) in all the ganglia studied. This explains that this particular stage of cellular growth (very-small cell stage) is a critical period during development, indication a stage of active cell-process formation (axon-formation), beginning to establish functional connections with the target tissues.
RESULTS
The results of cell-counts of different categories of cells (Fig: 1) are presented in the form of tables in order to avoid too many descriptions, for various ganglia as illustrated below.
| A. | DARK AND LIGHT CELLS (Please refer to the tables given above) In general, by a critical analysis and evaluation of the results in all the ganglia studied in the present series of investigation, it is assumed that |
| a. | the dark cells represent a group of functionally active cells which might proliferate, grow, mature, establish proper connections and continue to perform their functions. However, these cells may lose their activity and become inactive or die at any stage of their development, growth or activity and change to a light coloured cell on staining. |
| b. | the light cells represent a group of inactive, dying, dead or degenerating cells. In many situations, the occurrence of light cells in the ganglion for the first time is associated with loss of cells. These cells might become inactive or die due to some inherent defects developed within themselves or to some adverse factors found in the micro-environment. These light cells are found to appear around the time when the cells begin to establish their projections and represent those which fail to establish functional connections. However, sometimes when the adverse factors are rectified, these cells which have at first started to lose their functions might be re-activated and become normal active cells again and, therefore, might turn to be a dark type(Please refer to the results in ganglia where total-cells number and dark cells increase while there is no evidence of proliferation: by the occurrence of tiny and smaller categories of cells. This might be possible either. Results are showin in Table 1-9. |
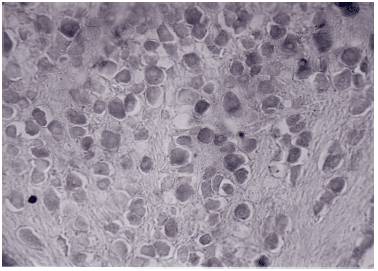 | |
| Fig: 1: | Shows dark neurons (with dark cytoplasmic stain) and light neurons (with light cytoplasmic stain) in the developing ganglion |
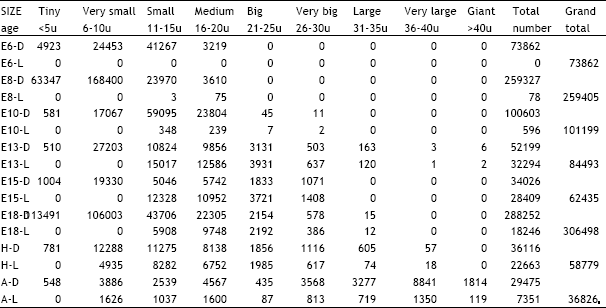
| Table 1: | Illustrates the total number of dark and light cells in the trigeminal ganglion in different age groups of animals in the ontogeny of the chick. D = Dark cells, L = Light cells, E = Embryonic age, H = Day of hatching, A = Adult |
 | |
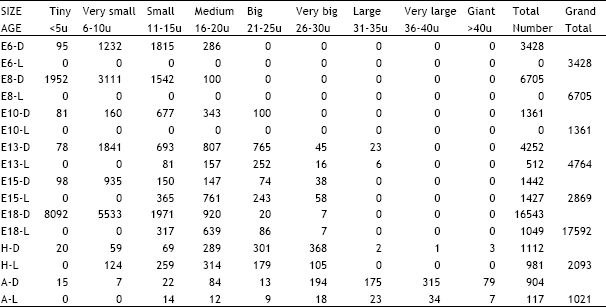
| Table 2: | Illustrates the total number of dark and light cells in the geniculate ganglion in different age groups of animals in the ontogeny of the chick D = Dark cells, L = Light cells, E = Embryonic age, H = Day of hatching, A = Adult |
 | |
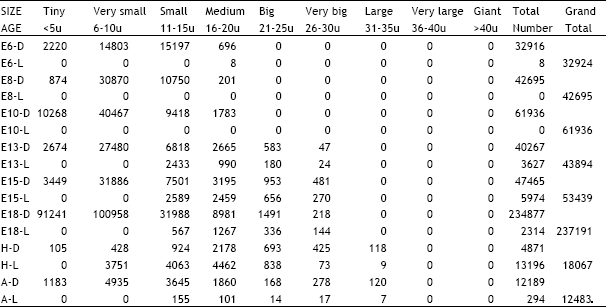
| Table 3: | Illustrates the total number of dark and light cells in the vestibular ganglion in different age groups of animals in the ontogeny of the chick D = Dark cells, L = Light cells, E = Embryonic age, H = Day of hatching, A = Adult |
 | |
| Table 4: | Illustrates the total number of dark and light cells in the acoustic ganglion in age groups of animals in the ontogeny of the chick D = Dark cells, L = Light cells, E = Embryonic age, H = Day of hatching, A = Adult |
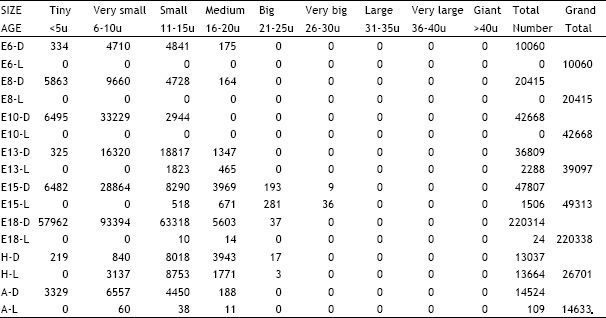 | |
| Table 5: | Illustrates the total number of dark and light cells in the proximal ganglionic complex of cranial nerves IX and X in different age groups of animals in the ontogeny of the chick. D = Dark cells, L = Light cells, E = Embryonic age, H = Day of hatching, A = Adult |
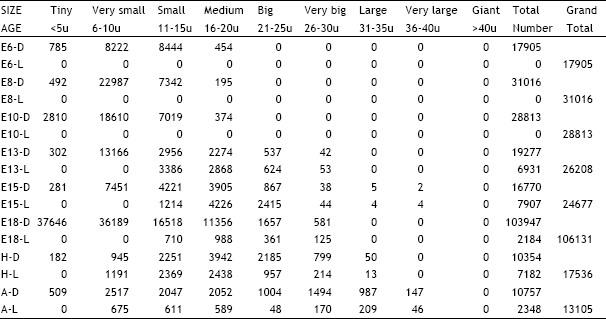 | |
| Table 6: | Illustrates the total number of dark and light cells in the petrous ganglion in different age groups of animals in the ontogeny of the chick D = Dark cells, L = Light cells, E = Embryonic age, H = Day of hatching, A = Adult |
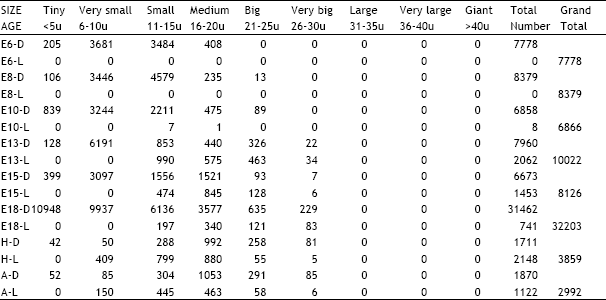 | |
| Table 7: | Illustrates the total number of dark and light cells in the nodose ganglion in different age groups of animals in the ontogeny of the chick D = Dark cells, L = Light cells, E = Embryonic age, H = Day of hatching, A = Adult |
 | |
| i. | by re-activation of their same original cell-processes which have first failed to establish a functional projection into their peripheral field of innervation, by rectifying the defects (found either within the cells themselves or in their micro-environment) by developing some favourable conditions, or |
| ii. | by the development of new collateral branches from the main process, which might grow new and establish functional connections to their innervation fields. |
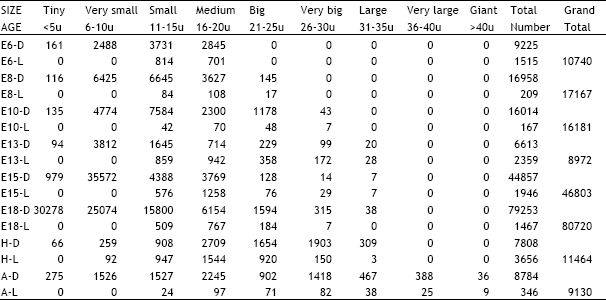
| Table 8: | Illustrates the total number of dark and light cells in the ciliary ganglion in different age groups of animals in the ontogeny of the chick D = Dark cells, L = Light cells, E = Embryonic age, H = Day of hatching, A = Adult |
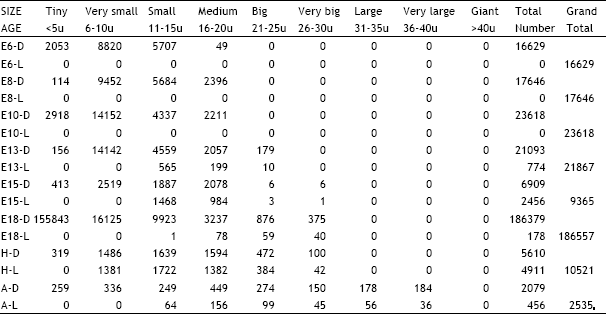 | |
| Table 9: | Illustrates the total number of dark and light cells in the superior cervical ganglion in different age groups of animals in the ontogeny of the chick. D = Dark cells, L = Light cells, E = Embryonic age, H = Day of hatching, A = Adult |
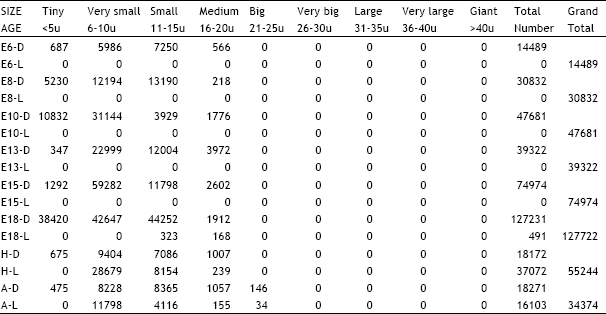 | |
The following evidences are presented in support of this assumption about the significance of the dark and light groups of cells. However, these statements are given in the form of brief points in order to avoid unnecessary long descriptions which might also need repetitions for every ganglion studied. Later, the facts are described in relation to some of the relevant available literature. Whenever necessary, for more details, the results of that particular ganglion may be verified and the facts be confirmed. The percentage or ratio of the dark and light cells in the ganglion in any age group may be found in the description of the results.
| 1. | During the periods of active proliferation and growth especially in the early stages of development, while there is a continuous increase in the total number of cells, all the cells among all categories found in the ganglion are dark type; no light cells are found during these active periods of development. Therefore, the dark cells are considered as a group of active cells which might divide, proliferate, grow, mature and thereby help to increase (or add) the number of all classes of cells in the ganglion. It may be noticed that the light cells begin to appear in the respective ganglia just after the stages given below. The time of appearance of light cells in the ganglion is presumed to be related to the time of their failure to establish functional connections. The beginning of establishment of such connections should be earlier than the time of occurrence of these light cells in the ganglion. Of course, this period varies from one ganglion to the other. The following description shows the developmental stages where all the ganglion cells are dark type after which the light cells begin to appear. |
| Trigeminal ganglion | E6, probably the establishment of functional connection begins early |
| Geniculate ganglion | E6, E8, E10 |
| Vestibular ganglion | E6, E8, E10; Please refer to the explanation given for the presence of a few (8) light cells on E6 |
| Acoustic ganglion | E6, E8, E10 |
| Prox. G. Comp. of IX and X | E6, E8, E10 |
| Petrous ganglion | E6, E8 |
| Nodose ganglion | Light cells are found even on E6; Probably the establishment of functional projections begins very early, even before E6. Thus this is the first ganglion to develop functional connections, possibly related to its supply to vital organs such as heart, lungs and aliment-ary canal and its importance. |
| Ciliary ganglion | E6, E8, E10 |
| Superior cervical ganglion | E6, E8, E10, E13, E15; Very late appearance of light cells |
Exceptionally, in certain stages of development, in some of these ganglia (given below), the number of cells are reduced instead of a continuous increase, indicating probably the presence of an active phagocytic process which would remove the inactive or dead cells (so-called light cells) during developmental period. However, the light-cell stage is not represented in them probably because the phagocytosis is so active and so-fast to leave this light-cell stage for clear observation. These developmental stages probably represent some critical periods (in that particular ganglion) in their attempt to establish functional projection to their innervation fields. It may be noticed that most of these periods of cellular loss are just before the occurrence of light cells in the ganglion.
| Geniculate ganglion | E10 |
| Prox. G. Comp. of N. IX and X | E10 |
| Superior cervical ganglion | E13 |
| 2. | As soon as the light cells have appeared in the ganglion, the total cells have reduced in number representing a loss of cells which, in turn, suggests that these light cells play a role in the loss or reduction of cells in the ganglion, or in other words, these light cells might represent a group of resting, inactive, dying, dead or degenerating cells which will, in course of time, be removed from the ganglion by phagocytes. It may be noticed that the loss or reduction in the total number of cells in the ganglion is occurring around the time when the light cells make their first appearance. |
| Trigeminal ganglion | E10, the very few (78 cells) negligible number of light cells found on E8 probably have formed just then, which is the beginning of cell death where the total cells however are greater in number. |
| Geniculate ganglion | E15, the few (512 cells) light cells found on E13 probably is the beginning of an observable cell death. The cell loss found on E10 is explained in the Discussion; probably the removal is too fast so as not to observe the light-cell stage. |
| Vestibular ganglion | E13 |
| Acoustic ganglion | E13 |
| Prox. G. Comp. of N. IX and X | E13 |
| Petrous ganglion | E10 |
| Nodose ganglion | Light cells are found even on E6; earlier stages are not observed in this ganglion. |
| Ciliary ganglion | E13 |
| Superior cervical ganglion | A few (491 cells = 0.38 %) light cells have appeared only on E18, which is probably the eginning of the appearance of light cells, however, on the day of hatching a gross reduction in the total number of cells having an increased number of light cells is found. However, in the earlier stages of cell loss (E13) light cells are not found. |
| 3. | On E18, there is usually a greatly reduced number of light cells (compared to E15) in relation to the tremendously increased number of dark cells (predominantly smaller categories), most of which are probably phagocytes (please refer to Part C : Removal of Dead Cells in the end) (also assumed from the present results found on the day of hatching where there is a great loss in the total number of cells in the ganglion). It is possible that the phagocytic activity is so great and so-fast that the light cell stage is not always observable since most of these inactive cells are actively removed from the vicinity of the ganglion before they become observable. This is a constant feature in all the ganglia in order to free the tissue from the noxious effects of the remnants of dead cells before the delicate and young animal is exposed to an independent living on the day of hatching. This is true in all the ganglia studied except a small difference observed in the superior cervical ganglion where the light cells have appeared for the first time only on E18. |
| 4. | a. But on the day of hatching, there is usually a greater proportion of light cells in the ganglion while there is a greatly reduced total number of cells because most of the unwanted cells have been removed by the greatly increased phagocytic activity found around E18. During post-hatching period, the light cells are greater in number. Probably, most of these light cells are in a temporary resting or inactive stage; many of them might become active functional cells again. It is also assumed that a proportion of the smaller categories of dark cells might represent the continued presence of phagocytes, ready to remove the inactive or dead cells. This is true in all the ganglia studied except Geniculate ganglion where some differences are found. That is, here in the geniculate ganglion greatest number of light cells are found on E15; later the light cells reduced in number throughout embryonic development, on the day of hatching as well as in the adult situation. |
| b.The tiny cells are found to be always dark. The very small type of cells are also found to be dark through the whole embryonic period till E18. Later, however, the light cells have appeared among the very small type also on the day of hatching, but they may or may not continue to be present in the adult situation. This might imply that even though the very small type of cells appear to keep themselves to be an active group till the day of hatching, and be ready to replace the dead cells occurring as a result of several adverse factors, cell death and degeneration begin among these cells also as from the day of hatching. It may be assumed that normally there cannot be any more necessity for the establishment of new functional projections after the day of hatching since all these connections might have been already established by this time while the animal is ready to lead an independent living. Therefore, there is no need for further growth and maturation of this smaller category of cells and the cell death begins even among this very small type of cells as from the day of hatching. Thus the appearance of light cells among this group just on the day of hatching is suggestive of evident cellular inactivity, death and degeneration process. |
| Trigeminal ganglion | True : Continue to be present in the adult |
| Geniculate ganglion | True : Disappear in the adult |
| Vestibular ganglion | True : Disappear in the adult |
| Acoustic ganglion | True : Continue to be present in the adult |
| Prox. G. Comp of IX and X | True : Continue to be present in the adult |
| Petrous ganglion | True : Continue to be present in the adult |
| Nodose ganglion | True : Disappear in the adult |
| Ciliary ganglion | True : Disappear in the adult |
| Superior cervical ganglion | True : Continue to be present in the adult |
| 5. | Even the larger classes of cells (having a diameter greater than 30 microns) whenever they have appeared in the ganglion contain both dark as well as light cells. (However, the dark and light cells are also found among smaller classes as well). This contradicts the descriptions of many earlier workers (Gaik and Farbman 1973, Noden 1978, Kishida et al 1985) that the ganglion contains small dark cells and large light cells and also contradicts their attribution of different functions to these cells because, in the present study the dark and light cells are found distributed among all categories whose diameter is greater than 10 microns irrespective of their small or large size. Therefore, such classification and functional attribution are disputed. |
| Trigeminal ganglion | E13, E18, on the day of hatching and adult; on E15 these larger classes of cells have totally disappeared |
| Geniculate ganglion | E13 and adult; on E15 and E18 the larger classes of cells have totally disappeared; on the day of hatching only dark cells are found. |
| Vestibular ganglion | On the day of hatching and adult when these larger classes of cells have appeared. |
| Acoustic ganglion | Such large class of cells has never appeared in the ganglion |
| Prox. G. Comp. of IX and X | E15, day of hatching and adult when the larger classes of cells have appeared. On E18 these larger classes have totally disappeared |
| Petrous ganglion | Such large class of cells has never appeared |
| Nodose ganglion | E13, E15, day of hatching and adult when these larger classes of cells have appeared. On E18 only dark cells are found among them. |
| Ciliary ganglion | In the adult, only when these larger classes of cells have appeared |
| Superior cervical ganglion | Such large class of cells has never appeared |
| 6. | The light cells continue to be present in the ganglion even in the adult situation while the total number of cells also continue to reduce. This is probably due to cellular inactivity and death as a result of ageing process while the functional reduction or functional loss is found in all organs including the organs of special sensibility and nervous control. This factor is uniformly noticed in all the ganglia studied. |
| 7. | The number of light cells lost in the ganglion is almost equal to the loss in the total number of cells, at a stage while the proliferation has stopped or reduced as evidenced by the number of tiny cells. Even though such condition is observed in a few instances in the whole investigation, this cannot be neglected as invalid because such incidence or circumstance cannot be expected to occur frequently in a constantly changing life cycle in the ontogeny, when such a change has to coincide with the time of observation. |
| Petrous ganglion | Compare the results of these ganglia on the day of hatching and the adult situation, at a time while the tiny cells are almost equal in number that indicates the stoppage of proliferative activity |
| Superior cervical ganglion | “Compare the results of these ganglia on the day of hatching and the adult situation, at a time while the tiny cells are almost equal in number that indicates the stoppage of proliferative activity |
| 8. | In the early stages of development, only dark cells are found in all the ganglia studied in the present series of investigation indicating that these are active cells. The light cells appear only after certain period of embryonic growth, probably at a time when the cells fail to establish proper functional projection onto their innervation fields. For example, in the present study, the structural evidence of the occurrence of light cells in the trigeminal ganglion on E8 seems to coincide with the appearance of physiological evidence occurring on the same embryonic day (E8) of exhibiting reflexogenic response to tactile stimulus of the beak of the chick embryo (Hamburger and Narayanan 1969). Similarly there are suggestions (Noden 1980, Gaik and Farbman 1873) that the placode-derived neurones in the trigeminal ganglion have well-established peripheral projections by the end of the first week of incubation. In contrast, the neurones derived from the neural crest in the trigeminal ganglion (Yates 1961, D=Amico Martel and Noden 1980) do not cease dividing until the seventh day of incubation. It may be recalled that the process-formation of neurones begins after the terminal mitosis which is considered as neurone=s birth date (D=Amico Martel and Noden 1980). Even though there is a slight variation in these suggestions given by different workers, it may be assumed that such degeneration is quite likely to happen around the seventh day of incubation. Thus the coincidence of appearance of light cells in the trigeminal ganglion in the present study indicating cell death on their failure to form proper connections suits very well with all these descriptions. However, similar physiological observations and reports for other ganglia are not available in the literature in order to make a comparison with the results in other ganglia studied in the present series of investigation. |
| 9. | It may also noticed that the light cells appear in the ganglion for the first time usually among small and medium sized cells (having a diameter of 11-20 microns) and later once they grow into larger classes, the light cells continue to be present among them. This is suggestive that the establishment of functional projection begins during this stage and that once they are unable to make functional connections, these cells become inactive or die and change into light coloured cells on staining. Such light cells may represent cellular inactivity or death. The light cells among larger classes may also represent cellular inactivity or death during successive growth periods, before establishing functional connections which may be due to any defect, either within the cell, in the micro-environment, inadequate supply of nerve growth factor (neurotrophic factor), or to ageing process in the case of adult. |
| 10. | However, it may be thought that the period of active cell loss (death) coincides with the period of active establishment of functional connections and in the present study, this period varies from ganglion to ganglion. This may be judged by the time of occurrence of light cells for the first time in the ganglion. However, this active establishment of functional connections may extend for longer periods as indicated by greater cellular loss in other stages. The following stages show the time of first appearance of light cells in the ganglion. |
| Trigeminal ganglion | E8 |
| Geniculate ganglion | E13 |
| Vestibular ganglion | E6, even though light cells are missing on E8 and E10 |
| Acoustic ganglion | E13 |
| Prox. G. Comp. of N. IX and XE | 13 |
| Petrous ganglion | E10 |
| Nodose ganglion | E6, early and frank appearance of light cells |
| Ciliary ganglion | E13 |
| Superior cervical ganglion | E18 |
The descriptions of the dark and light neurones by different investigators in different animals vary greatly in available literature. Some of these conflicting views from the literature which are considered more useful are described below. Some investigators (Peach 1972, Gaik and Farbman 1973) have described in the trigeminal ganglion of the chick that the large neurones contain many isolated clumps of basophilic material in a neurofilament-rich cytoplasmic matrix having a diameter ranging from 19 - 40 microns and that the smaller neurones having a diameter ranging from 11 - 30 microns tend to be multipolar and have a greater concentration of ribosomes and granular endoplasmic reticulum which are dispersed through a dense matrix. Based on their staining properties, these larger ones are called light neurones and the smaller ones are called dark neurones. In the adult rats, trigeminal neurones have been classified into dark and light types on the basis of distribution of cytoplasmic organelles within the neurones and on the relative density of cytoplasm (Dixon 1963, Carmel and Stein 1969, Matsura et al 1969). In a comparative ultra-structural study of the trigeminal ganglion (Moses et al 1965) small dark cells were densely packed with rough endoplasmic reticulum and ribosomes, and large light cells with a dispersed and occasionally clumped ergastoplasm. It may be noticed that the classification (Peach 1972, Gaik and Farbman 1973) of dark (small) and light (large) neurones overlap in size. However, all the above investigators have found that the dark (smaller) neurones have a greater concentration of ribosomes and granular endoplasmic reticulum, which is suggestive of a more active role than that of their lighter (larger) counterparts. This observation also supports the present results that the dark cells represent a group of active cells and the light cells represent a group of resting, inactive, dying, dead or degenerating cells.
However, in subsequent reports on human autopsied trigeminal ganglia, it was not easy for these workers (Moses 1967) to readily identify dark and light neurones. The majority of cells were intermediate in appearance. On rare occasions when they did observe these two neuronal types, the cells differed only in their cytoplasmic density and not in the arrangement of any of their organelles. These observations led these investigators to conclude that the dark and light cells were not real entities, but resulted from fluid-shifts between the cells in question and the surrounding extra-cellular spaces. By the same token, other investigators (Pineda et al 1967, Maxwell 1967) working with cat and monkey trigeminal ganglia found that if proper tissue fixation techniques were used, there were no base for classifying the trigeminal neurones into dark and light cell types. It is possible that these conflicting reports given by different investigators may be attributed to the species difference of the experimental animals used.
There has been suggestions (Gaik and Farbman 1973) that shortly before hatching (E18 onwards), there appears two classes of interposed neurones characteristic of the mature trigeminal ganglion : large light types and small dark type. Neither of these two populations corresponds uniquely to either of the two segregated populations (large peripherally and distally located cells, and small centrally and proximally located groups) found in the embryo. In the present study in the chick, the dual cytology of neurones (dark and light cells) is found in all the ganglia studied. They were observed not only in the mature ganglia (from 18th day of incubation to adult) as stated by earlier investigators (Peach 1972, Gaik and Farbman 1973, Noden 1978, Yates 1961, Hess 1955, Finkelbrand and Siberman 1977), but also in earlier developmental stages (please refer to the results). It is not clear whether the above workers failed to observe these classes of cells in earlier developmental stages because of unreliable staining techniques or whether they did not attempt to investigate their presence during early embryonic periods. It is also noticed in the present investigation that in the early stages of development, only dark cells are found in all the ganglia. The light cells begin to appear only after certain period of embryonic growth which varies from ganglion to ganglion : probably the time of their occurrence coincides with their failure to establish proper functional projection to their innervation fields. For example, in the present study in the trigeminal ganglion, the light cells have appeared for the first time on E8 which seems to coincide with the day (E8) of establishment of functional connections as observed on the basis of the presence of reflexogenic responses to tactile stimulus of the beak (Hamburger and Narayanan 1969).
It is frequently suggested (Hamburger 1961, Gaik and Farbman 1973, Noden 1978, Yates 1961, D= Amico-Martel and Noden 1980, Johnston 1966, Noden 1975) that the large light and small dark neurones found in the trigeminal ganglion are correlated with the dual embryonic origin as derived from both the epidermal placode and neural crest. Through most of the second week of development the large cells are found in the distal and ventro-lateral parts of the ganglion and the small cells are found in the proximal (core) and medio-dorsal parts (Hamburger 1962, Ebendal and Headlund 1975). Even though the present study is not aimed at finding out the embryological origin of these two categories of cells, the segregation of large cells in the distal and ventro-lateral parts and the small cells in the proximal (core) and medio-dorsal parts of the ganglion is evident from the present results similar to these findings.
Also there is both circumstantial and direct evidence that cytological dichotomy in the adult ganglion is not based on separate embryological origin. First, other sensory ganglia that are exclusively of neural crest origin such as trunk dorsal root ganglia, or of placodal origin such as acoustic ganglion have both dark and light types of neurones in the present study. Similar views have been advocated by earlier investigators (Rosenbluth 1962, Spoendlin 1974) also. In addition, the light and dark neurones are found interspersed at random throughout the ganglion during embryonic development and post-hatching periods in the present study, whereas the segregation of crest and placode-derived neurones (small dark and large light cells respectively) are found only in the embryo. Both transplantation experiments (Noden 1978) and birth-date analysis (D= Amico-Martel and Noden 1980) have proved that neurones from each of these anlagen retain their original separate locations during later stages of development and maturation. Thus the dual cytology of the mature trigeminal ganglion is not based on separate embryonic origins.
Since there is no clear evidence of trigeminal neuron projection to the solitary nuclear complex that normally receives only visceral input (Dubbeldam 1980, Dubbeldam and Karten 1978, Arends and Dubbeldam 1984), there should be no visceral neurones in the trigeminal ganglion. The work on trigeminal ganglion (Kishida et al 1985) have demonstrated that the trigeminal ganglion had the smallest proportion of dark cells (46.8 %) whereas in the proximal ganglionic complex of the cranial nerves IX and X and the distal ganglion of cranial nerve IX the proportion of the dark cells was much greater (84.7 % and 70.3 % respectively). Since these two ganglia have both somatic and visceral neurones (Dubbeldam et al 1979) the large proportion of small dark cells suggests that at least some of the dark cells are visceral neurones. In a work on spinal ganglia (Cervero et al 1984), the visceral neurones had a tendency to be smaller than somatic neurones. If this is true, then the larger portion of dark cells in the proximal ganglionic complex of cranial nerves IX and X observed by the above workers suggests that this ganglion has more visceral neurones than the distal ganglion of cranial nerve IX. The specific functional attribution for the neurones, based only on size difference (small and large cells) or staining properties (dark and light cells) as suggested by the above-mentioned workers, however, could not be confirmed in the present study because of the following reasons:
| a) | In the present investigation in the chick in the adult situation, even though the proportion of dark cells in the proximal ganglionic complex of cranial nerves IX and X (82.08 %) and that of the petrous ganglion of cranial nerve IX (62.5 %) are very close to the observations of the above mentioned workers in the same ganglia, the proportion of the dark cells found in the trigeminal ganglion (80.04 %) in the present study is quite different from that (46.8 %) observed by these investigators. This suggests that such proportion of dark and light cells in the ganglion might not be a constant factor so as to generalise their functional significance as has been suggested by them, or such changes could be due to species difference of the experimental animals as well. |
| b) | In the above-mentioned ganglia as well as in other ganglia studied in the present series of investigation, the proportion of dark and light cells shows a constant fluctuation through the whole ontogeny, i.e., during embryonic period, on the day of hatching as well as in the adult situation. Therefore, similar observation done at any one stage of the whole life cycle cannot prove valuable or give concrete evidence for such functional attribution for any cell. |
| c) | Again, this proportion of dark and light cells in the same ganglion is strikingly different in the adult situation even that observed on the day of hatching. The fluctuations in the number of these cells observed during development are considered as a necessary change for the establishment of a suitable functional organization in the animal. However, similar suggestion cannot be attributed to an animal on the day of hatching, a stage while the animal is already prepared for an independent living (just as the adult animals themselves). |
| d) | In the adult situation, in all the ganglia studied in the present series of investigation, there is not only a change in the proportion of dark and light cells but also a great reduction in the total number of cells in the ganglion. Similar variation again confirms that such method of functional attribution of cells, that too found just on one stage of a long life-cycle, and just on the basis of their staining characteristics cannot be validated because this staining characteristics may change in different conditions such as change in pH and functional state of that particular cell. Thus, on analysing the results in the present series of investigation, it is assumed that the light cells represent a group of resting, inactive, dying, dead or degenerating cells and the dark cells represent a group of functionally active cells in the ganglion. It is also assumed that the time of appearance of light cells in the ganglion is directly related to the onset of establishment of functional connections whose importance is related to the organs which their fibres supply, for example, early appearance of light cells in the nodose ganglion is probably related to the functional importance of the vagus nerve which supplies the vital organs such as the heart, lungs and alimentary canal which should be properly innervated as early as possible. |
It is also thought useful to quote the views of a few earlier investigators on Cell Death and Degeneration, and Removal of Dead Cells in order to complete the full evaluation of the dark and light cells. In this view, the following points are given for relevant reference.
Cell death and degeneration
One feature of development of many parts of the nervous system is the occurrence of two opposing processes: cellular proliferation which leads to the production of large numbers of neurones and massive cellular degeneration which results in the loss of many of these same neurones. These two processes ultimately control the final number of neurones of a neural centre. Even though no attempt was taken to investigate the purpose, reasons or ways of causing cell death which occur in different ganglia studied in the present series of investigation, the suggestions of some of the earlier investigators which are found suitable are given below in order to supplement the present assumption derived from a critical analysis of all these results. In the present study, cell death occurs mainly around E10 - E13 after which the ganglion prepares itself for a massive phagocytic activity which nearly comes to an end by the day of hatching. This resembles the descriptions (Hamburger 1934) that there are corresponding and parallel changes in the brain stem auditory nuclei and their peripheral ganglionic projections.
The observations (Hamburger 1934, 1958, Levi-Montalcini 1949, Cowan 1973) support the idea that it is the events in the target tissue (i.e., the periphery) which normally regulate cell survival. They demonstrated that the periphery both controls the proliferation and initial differentiation of undifferentiated cells and also provides the conditions necessary for continued growth and maintenance of neurones in stages following the first outgrowth of neurites. Several experiments (Hamburger 1958, Hamburger and Levi-Montalcini 1949, 1975, Prestige 1967, 1976, Kelly and Cowan 1972, Landmesser and Cowan 1974, Sohal 1976) have led to a general acceptance of the idea that neuronal death regulates nerve cell numbers in response to peripheral demands by eliminating those nerve cells whose fibres fail to establish proper peripheral connections. There are evidences (Cowan 1973, Hamburger 1975, Prestige 1970) suggesting that events at the target tissue controlling normal cell-death, involve the notion of either a competition between neurones for a limited number of synaptic sites and / or for a limited amount of trophic substances supplied by the target. Failure of the neurones to either make or receive the appropriate synaptic connections has been attributed to most neuronal deaths during embryogenesis (Cowan 1971, 1973). It also seems more likely that the main function of cell death in this system is probably to remove redundant neurones which though being in the correct muscle, have failed to form a contact (Landmesser and Morris 1975). According to this argument, the nervous system is programmed to over-produce cells in order to saturate the target and insure that all muscle fibers become innervated. The cell death presumably plays a role in the normal formation of orderly connections.
In some experiments (Hamburger and Levi-Montalcini 1949, Prestige 1970) in which it has been possible to experimentally enlarge the projection fields of the neuronal population, or to increase the number of afferents which it receives (a technique known to experimental neuro-embryologists as “ peripheral over-loading or peripheral enlargement”) the number of neurones found in the population at later stages of development has been significantly greater than normal. In contrast to these experiments, peripheral ablation experiments (Hamburger 1934, Kelly and Cowan 1972, Landmesser and Pilar 1974, Prestige 1970) have shown that the severity of the normal neuronal degeneration is much increased resulting in the additional cell death usually occurring over the same period as the naturally occurring neuronal loss. That is, natural cell death is known to be greatly enhanced by peripheral depletion.
From the available evidence, it is convincing to accept the idea that the peripheral influences, peripheral demands, trophic factors supplied by the target organs etc are important factors in controlling the neuronal death and the number of functional neurones available in the ganglia and that the neuronal death plays a role in the normal formation of orderly connections.
Removal of dead cells
Several investigators have suggested that macrophages are important in removing debris of dead cells (Levi-Montalcini 1950). Different sources of these macrophages have been suggested. One possibility is the transformation of mononuclear leucocytes in the circulatory system into tissue macrophages (Marchasi 1964, Sulton and Weiss 1966, Furth and Cohn 1968, Kitamura et al 1972). It has also been demonstrated (Adrian and Smotherman 1970) many radioactively labelled mononuclear leukocytes can infiltrate into nervous system and aggregate around damaged nervous tissue. There is also electron-microscopic evidence that the leukocytes invade nervous system by crossing through the wall of the blood capillaries (Matthews and Kruger 1973). It has also been demonstrated in an ultra-structural study (Wang-Chu and Oppenheim 1978) that the phagocytic cells contain neuronal debris that exhibit most of the characteristics of mononuclear leukocytes. The phagocytic activity of the satellite cells in the chick embryonic spinal ganglia are attributed to the removal of cellular debris of degenerated cell (Tennyson 1970) during early development. There are also reports (O=Connor and Wyttenbach 1974, Pilar and Landmesser 1976) that phagocytosis is accomplished by glial cells. However, some investigators (Michaels et al 1971) believe that the degenerating cells produce hydrolytic enzymes for their own degeneration. Therefore, there is obvious reason to believe that the leukocytes from the blood stream can penetrate through the wall of the capillaries into the nervous system and function as tissue macrophages to remove the neuronal debris during cell death and that the glial and satellite cells also can act as phagocytes.
Research summary
The chicks, Gallus gallus domesticus, White Leghorn breed were used in this study. The fertilised eggs were incubated at a temperature of 37.5 degree Centigrade. Embryos from the 3rd embryonic day onwards up to hatching were taken carefully without causing damage. They were immediately fixed in 10% formaldehyde solution and kept in the fixative for at least two weeks. The pieces having the head, neck and thorax were processed separate and paraffin blocks were prepared. The head of the adult animal rostral to the level of the 6th cervical vertebrate was separated and fixed in the fixative. Wherever necessary, the tissues were decalcified after fixation. The nodose ganglion in the adult was dissected out carefully fixed in the fixative and processed like other tissues. Serial sections having a thickness ranging from 8-10 microns were taken and stained by cresyl fast violet. In the embryos of 3-5 days, even though the ganglionic configuration of the different ganglia was noticed, their exact identification could not be done with accuracy. Therefore, the present study will include only the following stages of the chick which are of value of some critical interest in one ganglion or the other - E6, E8, E10, E13, E15, E18, The day of hatching, and the adult. The ganglia used for observation are ciliary, trigeminal, geniculate, vestibular, acoustic, superior ganglionic complex of glossopharyngeal and vagus nerves, petrous, nodose, superior cervical and 4th cervical spinal ganglia. Different types of cell were classified according to the intensity of stain in the cytoplasm and their size. Counting and categorizing the different types of cell were based on the camera lucida drawings of individual sections. Wherever possible, immunocyto-chemistry and microwave techniques were employed. The behaviour of different cell groups supposedly originating from the epidermal placodes and the neural crest were followed.
On the evidence available, the dark cells are considered as active ones; the light cells are considered as those which have failed to establish proper functional projections and are therefore destined to die and disappear from the vicinity of the ganglion and are classified as inactive, dying, dead or degenerating ones.
Even though the cell death and degeneration take place during all stages of cellular growth and maturation, it is most prominent and common among the small and medium sized one. Therefore, the small and medium sized ones. Therefore, the small and medium sized cells appear to be critical ones in the development of the ganglion; probably it is during these stages of cell growth the peripheral and central processes begin to grow from the cell body and attempt to get established in their projection fields. If they succeed in their attempt they continue to function and remain as dark cells, but if they fail in this attempt they lose their activity, tend to die and disappear, and change to a light coloured cell on staining. It is quite possible that some cells might lose their capacity and become light cells even after their processes grow into larger classes.
Since the small and medium sized cells are the youngest stages in which the light cells make their first appearance, it might indirectly mean that the establishment of connections might begin immediately after the very-small-celled stage, i.e., around small-cell stage. Successive larger classes of cells might represent the degree of development and maturation of their processes and continuation of functional capacity of the cell. However, the inherent nature of the cells also forms a basis for them to grow larger in size.
The cells which have started to die or degenerate and change into light cells might regain their capacity under favourable conditions and turn into dark active cells again. This might add to the existing number of dark cells in the ganglion. This may be possible either by recovery of the defects which have earlier developed either within the cells themselves or in the micro environment or by developing new collateral branch in order to substitute or supplement the previous functional loss. Such behaviour is frequently observed in the vestibular ganglion and also occasionally in some developmental stages in other ganglia.
It is presumed that the cell death plays a role in the normal formation of orderly connections.
The tiny cells are always found to be dark type. At no time light cells are found among them.
The very small type of cells are also usually dark during embryonic development till E18. The light cells have appeared among the very small type of cells also just on the day of hatching which may or may not continue in the adult situation. This probably signifies the possible attempt to eliminate the growing cells since they are no longer needed to replace larger categories of cells which have already well-developed neuronal connections at this stage by the day of hatching.
The cellular proliferation in the early stages of development might concern mainly with neuronal elements even though a proportion of these cells might be phagocytes being necessary to eliminate dead cells at every stage of development, and those formed around E15 - E18 might be concerned with mainly phagocytes which help remove the unsuccessful or redundant neuronal elements which probably fail to establish (to give or receive) proper functional connections.
It is assumed that the phagocytic cells might develop either from macrophages, glial cells, satellite cells and/or blood borne leukocytes.
Even after hatching, a few cells might continue to have the potentiality to act as phagocytes, to be protective for the ganglion even in later stages of life.
It is reasonable to presume that the optimum number of functional neurons must have been formed by the day of hatching since the animal is prepared and ready for an independent living by then.
From the evidences available, it is possible to presume that the optimum number of functional neurons and connections must have been established even before E18.
Usually the number of light cells are reduced very much on E18 even though the cell population is the highest in the whole ontogeny indicating probably a stage of faster and active removal of dead cells by the tremendously increased phagocytic cells. The phagocytosis is too fast that the inactive, dying or dead cells (i.e., so-called light cells) are removed immediately as soon as they are formed, to leave such a light celled stage for clear observation.
The light cell stage becomes clearly observable only when the phagocytic process is slow and become prominent at a time when some of the important connections are being actively established.
The volume of the ganglion is the greatest on E18 during the whole embryonic period as observed in all the ganglia studied. However, the ganglion shows its greatest volume in the adult situation in the whole ontogeny except the vestibular and acoustic ganglia. In most ganglia, the volume shows a reduction before and after E18 (in which the ganglion is larger in size, and has a tremendously increased phagocytic activity) i.e., around E15 and on the day of hatching.
The behaviour of cells in the vestibular ganglion as well as in the acoustic ganglion appears to be nearly similar in many respects.
Volume of the adult ganglion is lesser than that found on E18 even though its rostrocaudal length is greatest in the adult situation.
There is the appearance of larger proportion of light cells on the day of hatching while these cells are reduced in the adult situation.
There is an increased number of tiny cells in the adult ganglion than that seen on the day of hatching, probably indicating the importance of a renewal system of older cells which becomes functionally weak, in order to keep a balanced function.
However, the appearance of a few light cells in the vestibular ganglion on E6 might indirectly indicate the beginning of an early establishment of a functional connection, since the light cells are presumed to show up once these cells fail to form proper functional projection.
In the vestibular ganglion on the day of hatching the number of light cells is more than that of the dark cells, while in the adult situation the reverse is the case. In the adult, the proportion of these cells changes where the larger proportion is dark and smaller proportion is light types; probably among the enormous number of light cells formed on the day of hatching which are presumed to undergo death and degeneration, the major portion has recovered from this tendency possibly gaining the normal activity and function again by a process of re-activation of the original fibres before it is too late or by the development and establishment of new collateral branches which becomes functional and substitute the original unsuccessful ones.
In the vestibular ganglion the appearance of an increased number of tiny cells in the adult animal is comparison to that seen on the day of hatching (different and peculiar from other ganglia) probably provides a chance to replace or substitute the inactive (or dead) cells during ageing process by their growth, maturation and establishing new functional connections in view of the importance of an efficient vestibular function for normal behaviour of the animal.
The acoustic ganglion is also peculiar in that the larger classes of cells which have appeared in several stages following E6 have at last have disappeared in the adult situation, leaving a condition similar to that observed in the embryo as early as E6, whereas in most ganglia studied the larger classes of cells appear a new as the age increases. Probably the larger classes of cells are functionally unimportant in this ganglion and are eliminated in the adult situation.
The acoustic neurones are more sensitive to damage by ageing process compared to the cells in other ganglia studied and the functional reduction might become more prominent by increasing age. A renewal system to replace older cells by growth of new tiny cells might be in action, peculiar and similar to the vestibular ganglion.
The cell population and its density in most ganglia become low in the adult situation.
The rostrocaudal length of the ganglion is usually the greatest in the adult situation in the whole ontogeny except the vestibular and acoustic ganglia.
On the day of hatching there is always a reduction in the total number of cells than that found on E18 but having an increased number and increased proportion of light cells. The volume of the ganglion is also usually found to be reduced during this period.
In the adult situation the trigeminal ganglion is the largest one among all cranial nerve ganglia. The next one, greater in size is the nodose ganglion of the vagus nerve.
The behaviour of cells in the genicular ganglion also appears to be peculiar by being irregular in that they do not follow any of the clearly defined formula or regulation which are seen in other ganglia.
At least in some situation the nerve cells are found distributed in the nerve trunk in longitudinal rows between the nerve fibres, e.g., facial and vestibulocochlear nerves.
While a fluctuation in the cell population is taking place in the ganglion during development and growth, there are usually one, two or three stages which show population increase through the whole ontogeny as described below.
Cellular population of the 4th cervical spinal ganglion has only one raise, i.e., on E18 and a single decrease on the day of hatching and later a further reduction of cells in the adulthood.
Some ganglia have two raises in the cell population - one on E8 and another on E18 and a reduction on the day of hatching and later a continued reduction in the adult situation. e.g., trigeminal ganglion, proximal ganglionic complex of nerve IX and X, and nodose ganglion.
Some ganglia have the first population raise on E10 and another on E18 and later reduces on the day of hatching and also in the adult. e.g., cilliary ganglion, vestibular ganglion, acoustic ganglion and superior cervical ganglion.
Still others have three periods of population raise, on E8, E13 and E18 and later reduces on the day of hatching and also in the adult. e.g., genicular ganglion and petrous ganglion.
The nodose ganglion is the only ganglion which contains frank light cells even as early as E6. This indicates that the extensive establishment of connections takes place even before this stage, probably due to its functional importance supplying the vital organs such as heart, lungs and alimentary canal. Similarly the vestibular ganglion also shows a few light cells on E6 even though no light cells are observed on E8, E10 etc which also probably implies the importance of vestibular function.
It may be thought that the time of appearance of light cells in the ganglion might be indirectly related to the onset of establishment of active functional connections and to the functional importance of the organs which it supplies.
The increased volume of the ganglion on E18 might be due to a combined effect of tissue reaction resulted from the toxic substances of dead cells along with the phagocytic activity which are necessary to protect the young and delicate animal from the toxic effects.
The cell-space available for every cell in the ganglion is the greatest in the adult situation while the cell number is greatly reduced and the ganglionic volume is greatly increased.
The reduction or loss in the number of neurones in the adult ganglion in comparison to that found on the day of hatching might indicate a functional reduction of the ganglion probably as a result of ageing process.
The fluctuations in the number of cells during embryonic development may be considered as a normal process for the purpose of re-arrangement and better organization in order to perform an efficient function.
It is seen that there appears to be an initial small increase in the volume of the ganglion approximately upto E10, then there is a fall during the period of rapid cell loss around E8-E15 and then a rapid increase around E13-E18. However, these periods vary from ganglion to ganglion. The major period of cellular degeneration is usually around E11-E15 of incubation even though its intensity, amount and duration differ in different ganglia. It is also noteworthy that cell loss has essentially ended around E15 by the time the ganglion begins to show the greatest expansion so as to reach its maximum size on E18 during embryonic development.
It is assumed that the period of accelerated degeneration (or cell loss) is the period of active establishment of proper connections of the ganglion cells.
There is usually a great increase in the volume of the ganglion between E15 of incubation and the day of hatching which is due in part to the increased size of the cell bodies, and to the increased size of the neuropil lying between them because of the development of a network of nerve fibres and blood vessels.
It may be agreed that several convergent intrinsic and extrinsic factors might play a role in the regulative and/or stimulative control of behaviour of the ganglion and its cells during proliferation, aggregation of cells into ganglion, growth, maturation, establishment of functional projections, cell death, degeneration, disintegration, removal of dead cells and ageing processes.
REFERENCES
- Arends, J.A. and J.L. Dubbeldam, 1984. The subnuclei and primary afferents of the descending trigeminal system in the mallard (Anas platyrhynchos L.). Neuroscience, 13: 781-791.
PubMed - Bondareff, W., 1987. Changes in the brain in aging and Alzheimers disease assessed by neuronal counts. Neurobiol Aging., 8: 562-563.
CrossRef - Cammermeyer, J., 1962. An evaluation of the significance of the dark neuron. Ergeb. Anat. Entwicklungsgesch., 36: 1-61.
PubMedDirect Link - Carmel, P.W. and B.M. Stein, 1969. Cell changes in sensory ganglia following proximal and disal nerve section in the monkey. J. Comp. Neurol., 135: 145-165.
CrossRefDirect Link - Coleman, P.D. and D. Flood, 1987. Neuron number and dendritic extent in normal ageing and Alzheimers disease. Neurobiol Aging., 8: 521-545.
PubMed - DAmico-Martel, A. and D.M. Noden, 1980. An autoradiographic analysis of the development of the chick trigeminal ganglion. J. Embryol. Exp. Morphol., 55: 167-182.
PubMed - Dixon, A.D., 1963. Fine structure of nerve cell bodies and satellite cells in the trigeminal ganglion. J. Dent. Res., 42: 990-999.
Direct Link - Dlugos, C.A. and R.J. Pentney, 1994. Morphometric analyses of purkinje and granule cells in aging F344 rats. Neurobiol Aging., 15: 435-440.
CrossRefDirect Link - Dubbeldam, J.L. and H.J. Karten, 1978. The trigeminal system in the pigeon (columba livia). Projections of the gasserian ganglion. J. Comp. Neurol., 180: 661-678.
PubMed - Dubbeldam, J.L., E.R. Brus, S.B.J. Menken and S. Zeilstra, 1979. The central projections of the glossopharyngeal and vagus ganglia in the mallard, Anas platyrhynchos L. J. Comp. Neurol., 183: 149-168.
PubMed - Finkelbrand, S. and M. Silberman, 1976. Effect of long-term hyperglucocorticoidism on the neuronal morphology of the trigeminal ganglion of the mouse. Acta Anatom., 96: 582-590.
CrossRefDirect Link - Flood, D.C. and P.D. Coleman, 1988. Neuron numbers and sizes in aging brain: Comparison of human, monkey and rodent data. Neurobiol. Aging., 9: 453-463.
PubMed - Furth, R. and Z.A. Cohn, 1986. The origin and kinetics of mononuclear phagocytes. J. Exp. Med., 128: 415-435.
Direct Link - Gaik, G.C. and A.I. Farbman, 1973. The chicken trigeminal ganglion I. An anatomical analysis of the neuron types in the adult. J. Morphol., 141: 43-55.
CrossRefDirect Link - Gobel, S., 1974. Synaptic organization of the substantia gelatinosa glomeruli in the spinal trigeminal nucleus of the adult cat. J. Neurocytol., 3: 219-243.
CrossRefDirect Link - Hamburger, V., 1934. The effects of wing bud extirpation on the development of the central nervous system in chick embryos. J. Exp. Zool., 68: 449-494.
CrossRef - Hamburger, V., 1958. Regression versus peripheral control of differentiation in motor hypoplasia. Am. J. Anat., 102: 365-409.
CrossRefDirect Link - Hamburger, V., 1961. Experimental analysis of the dual origin of the trigeminal ganglion in the chick embryo. J. Exp. Zool., 148: 91-123.
PubMed - Hamburger, V. and R. Levi-Montalcini, 1949. Proliferation, differentiation and degeneration in the spinal ganglia of the chick embryo under normal and experimental conditions. J. Exp. Zool., 111: 457-501.
CrossRefDirect Link - Hamburger, V., 1975. Cell death in the development of the lateral motor column of the chick embryo. J. Comp. Neurol., 160: 535-546.
CrossRefPubMedDirect Link - Hamburger, V. and C.H. Narayanan, 1969. Effects of the deafferentation of the trigeminal area on the motility of the chick embryo. J. Exp. Zool., 170: 411-426.
CrossRefDirect Link - Heinensen, H., H. Henn, W. Eisenmenger, M. Gotz and J. Bohl et al., 1994. Quantitative investigations on the human entorhinal area: Left-right asymmetry and age-related changes. Anat. Embryol., 190: 181-194.
CrossRefDirect Link - Hess, A., 1966. The fine structure of young and old spinal ganglia. Anat. Rec., 123: 399-423.
CrossRefDirect Link - Johnston, M.C., 1966. A radioautographic study of the migration and fate of cranial neural crest cells in the chick embryo. Anat. Rec., 156: 143-156.
CrossRef - Kalina, M. and J.J. Bubis, 1969. Ultrastructural localization of acetylcholine esterase in neurones of rat trigeminal ganglia. Experientia, 25: 388-389.
CrossRefDirect Link - Kalina, M. and M. Wolman, 1970. Correlative histochemical and morphological study on the maturation of sensory ganglion cells in the rat. Histochemie, 22: 100-108.
CrossRef - Kelly, J.P. and W.M. Cowan, 1972. Studies on the development of the chick optic tectum. III. Effects of early eye removal. Brain Res., 42: 263-288.
CrossRefDirect Link - Kishida, R., S. Terashima, R.C. Goris and T. Kusunoki, 1982. Infrared sensory neurons in the trigeminal ganglia of crotaline snakes: Transganglionic HRP transport. Brain Res., 241: 3-10.
CrossRefDirect Link - Kishida, R., J.L. Dubbeldam and R. Goris, 1985. Primary sensory ganglion cells projecting to the principal trigeminal nucleus in the Mallard, Anas platyrhynchos. J. Comp. Neurol., 240: 171-179.
CrossRef - Kitamura, T., H. Hattori, S. Fujita, 1972. Autoradiographic studies on histogenesis of brain macrophages in the mouse. J. Neuropath. Exp. Neurol., 31: 502-518.
PubMed - Levi-Montalcini, R., 1949. The development of the acoustico-vestibular centres in the chick embryo in the absence of the afferent root fibers and of descending fiber tracts. J. Comp. Neurol., 91: 209-241.
CrossRefDirect Link - Levi-Montalcini, R., 1950. The origin and development of the visceral in the spinal cord of the chick embryo. J. Morphol., 86: 253-283.
CrossRefDirect Link - Landmesser, L. and D.G. Morris, 1975. The development of functional innervation in the hind limb of the chick embryo. J. Physiol., 249: 301-326.
Direct Link - Landmesser, L. and G. Pilar, 1974. Synapse formation during embryogenesis on ganglion cells lacking a periphery. J. Physiol., 241: 715-736.
Direct Link - Marchasi, V.T., 1964. Some electron microscopic observations on interactions between leukocytes, platelets and endothelial cells in acute inflammation. Ann. N. Y. Acad. Sci., 116: 774-788.
CrossRefDirect Link - Matsura, H., M. Mori and K. Kawakatsu, 1969. A histochemical and electron-microscopic study of the trigeminal ganglion of the rat. Arch. Oral. Biol., 14: 1135-1146.
PubMedDirect Link - Matthews, M.A. and L. Kruger, 1973. Electron microscopy of non‐neuronal cellular changes accompanying neural degeneration in thalamic nuclei of the rabbit. I. Reactive hematogenous and perivascular elements within the basal lamina. J. Comp. Neurol., 148: 285-311.
CrossRefDirect Link - Maxwell, D.S., 1967. Fine structure of the normal trigeminal ganglion in the cat and monkey. J. Neurosurg., 26: 127-131.
PubMed - Michaels, J.E., J.T. Albright and D.I. Patt, 1971. Fine structural observations on cell death in the epidermis of the external gills of the larval frog, Rana pipiens. Am. J. Anat., 132: 301-317.
CrossRefDirect Link - Moses, H.L., 1967. Comparative fine structure of the Trigeminal ganglia, including human autopsy studies. J. Neurosurg., 26: 112-126.
CrossRefDirect Link - Moses, H.L., D.L. Beaver and C.E. Ganote, 1965. Electronmicroscopy of the trigeminal ganglion: Comparative ultrastructure. Arch. Pathol., 79: 541-556.
PubMed - Noden, D.M., 1980. Somatotopic and functional organization of the avian trigeminal ganglion: An HRP analysis in the hatchling chick. J. Comp. Neurol., 190: 405-428.
CrossRefDirect Link - O'Connor, T.M. and C.R. Wyttenbach, 1974. Cell death in the embryonic chick spinal cord. J. Cell Biol., 60: 448-459.
CrossRefDirect Link - Peach, R., 1972. Fine structural features of light and dark cells in the trigeminal ganglion of the rat. J. Neurocytol., 1: 151-160.
CrossRefDirect Link - Pilar, G. and L. Landmesser, 1976. Ultrastructural differences during embryonic cell death in normal and peripherally deprived ciliary ganglia. J. Cell Biol., 68: 339-356.
CrossRefDirect Link - Pillay, A.G., 1999. Acoustic ganglion in the chick during development and ageing process. Pak. J. Biol. Sci., 2: 1219-1223.
CrossRefDirect Link - Pillay, A.G., 1999. Developmental and age-related changes in morphometry and cellular density of the vestibular ganglion in the chick. Pak. J. Biol. Sci., 2: 1371-1374.
CrossRefDirect Link - Pillay, A.G. and G.D. Vennila, 1999. Organization, behaviour and significance of the changing pattern of neural elements in the trigeminal ganglion during development and aging: An investigation in the chick. J. Anat. Soc. Ind., 48: 73-82.
Direct Link - Pillay, A.G., 2000. Cell turn-over in trigeminal ganglion during gangliogenesis and aging process and its significance. Pak. J. Biol. Sci., 3: 1736-1739.
Direct Link - Pillay, A.G., 2000. Cellular death as a regulative mechanism in the control of cell-number towards normal establishment of orderly structure and function in the petrous ganglion during development and aging: An investigation in the chick. Pak. J. Biol. Sci., 3: 1743-1748.
CrossRefDirect Link - Pillay, A.G., 2001. Dual cytology, cellular death and their significance in the proximal ganglionic complex: An investigation through the ontogeny of the chick gallus gallus domesticus. J. Med. Sci., 1: 16-21.
CrossRefDirect Link - Pillay, A.G., 2001. Vagal and vestibular ganglia as vital neural-centers during development. J. Medical Sci., 1: 228-234.
CrossRefDirect Link - Pineda, A., D.S. Maxwel and L. Kruger, 1967. The fine structure of neurons and satellite cells in the trigeminal ganglion of cat and monkey. Am. J. Anat., 121: 461-487.
CrossRefDirect Link - Prestige, M.C., 1967. The control of cell number in the lumbar spinal ganglia during the development of Xenopus laevis tadpoles. J. Embryol. Exp. Morphol., 17: 453-471.
Direct Link - Prestige, M.C., 1967. The control of cell number in the lumbar ventral horns during the development of Xenopus laevis tadpoles. J. Embryo. Exp. Morpho., 18: 359-387.
Direct Link - Prestige, M.C., 1976. Evidence that at least some of the motor nerve cells that die during development have first made peripheral connections. J. Comp. Neurol., 170: 123-133.
PubMed - Rosenbluth, J., 1962. The fine structure of acoustic ganglia in the rat. J. Cell. Biol., 12: 329-359.
Direct Link - Silbermann, M. and S. Finkelbrand, 1978. Activity of phosphatases and succinic dehydrogenase in the Trigeminal ganglion of the corticosteroid-treated mouse. Acta Anat., 100: 229-236.
CrossRefDirect Link - Sohal, G.S., 1976. An experimental study of cell death in the developing trochlear nucleus. Exp. Neur., 51: 684-696.
PubMed - Spassova, I., 1982. Cat trigeminal ganglion. Neuron types. An experimental study. Z. Mikrosk. Anat. Forsch., 96: 235-244.
PubMedDirect Link - Sulton, J.S. and L. Weiss, 1966. Transformation of monocytes in tissue culture into macrophages, epithelioid cells and multinucleated giant cells: An electron microscopic study. J. Cell. Biol., 28: 303-332.
Direct Link - Tennyson, V.M., 1965. Electron microscopic study of the developing neuroblast of the dorsal root ganglion of the rabbit embryo. J. Comp. Neurol., 124: 267-317.
CrossRef - Tennyson, V.M., 1970. The fine structure of the axon and growth cone of the dorsal root neuroblast of the rabbit embryo. J. Cell Biol., 44: 62-79.
CrossRefPubMedDirect Link - Chu-Wang, I.W. and R.W. Oppenheim, 1978. Cell death of motoneurons in the chick embryo spinal cord. I. A light and electron microscopic study of naturally occurring and induced cell loss during development. J. Comp. Neurol., 177: 33-57.
CrossRefPubMedDirect Link - West, M.J., 1993. Regionally specific loss of neurons in the aging human hippocampus. Neurobiol. Aging., 14: 287-293.
PubMedDirect Link - Yates, R.D., 1961. A study of cell division in chick embryonic ganglion. J. Exp. Neur., 147: 167-181.
PubMed